
Enamik allpool olevast materjalist on avaldatud analüütilises aruandes. “Homoseksuaalse liikumise retoorika teaduslike faktide valguses”. doi:10.12731/978-5-907208-04-9, ISBN 978-5-907208-04-9
Peamised järeldused
1. Hüpoteetiline “homoseksuaalsuse geen” pole teada, seda pole keegi avastanud.
2. Homoseksuaalsuse sünnipärasuse väite aluseks olevad uuringud sisaldavad mitmeid metodoloogilisi ebatäpsusi ja vastuolusid ning ei võimalda teha selgeid järeldusi.
3. Isegi LGBT+ liikumise aktivistide viidatud olemasolevad uuringud ei räägi homoseksuaalsete kalduvuste geneetilisest määratlusest, vaid parimal juhul komplekssest mõjust, mille puhul oletatavasti määrab geneetiline tegur eelsoodumuse, kombineerituna keskkonnamõjude, kasvatusega, jne.
4. Mõned homoseksuaalliikumise prominentsed tegelased, sealhulgas teadlased, kritiseerivad väiteid homoseksuaalsuse bioloogilise ettemääratuse kohta ja väidavad, et see on põhjustatud teadlikust valikust.
5. LGBT propagandameetodite autorid «After The Ball» soovitas valetada homoseksuaalsuse sünnipärasuse kohta:
„Esiteks peab avalikkus olema veenev, et geid on asjaolude ohvrid ja et nad ei vali oma seksuaalset sättumust rohkem kui oma pikkust, nahavärvi, andeid või piiranguid. Hoolimata asjaolust, et ilmselt on enamiku inimeste seksuaalne sättumus keerukate koosmõjude tagajärg kaasasündinud eelsoodumuse ja keskkonnategurite vahel lapsepõlves ja varases noorukieas, nõuame kõigil praktilistel eesmärkidel, et geid on sündinud niimoodi.
<..>
Homoseksuaalid ei valinud midagi, keegi pole neid kunagi lollitanud ega võrgutanud.
Sissejuhatus
Argument, et homoseksuaalne külgetõmme on kaasasündinud - nn homoseksuaalse külgetõmbe bioloogilise determinismi hüpotees on LGBT + liikumise üks põhilisi. Loosung "Sel viisil sündinud"1, mida aktiivselt levitati popkultuuris, pani paljud mittespetsialistid arvama, et homoseksuaalsuse bioloogiline genees on midagi vaieldamatut ja tõestatud. See pole tõsi.
Kõige usaldusväärsemad faktid homoseksuaalsuse kohta ei viita mitte bioloogilisele, vaid sotsiaal-ökoloogilisele põhjuslikule seosele. Viimaste aastakümnete püüdlused leida andmeid, mis toetaksid bioloogilisi teooriaid, on vaid suurendanud kahtlust, kas sellised andmed on olemas.
Homoseksuaalsuse bioloogilise geneesi tees pole iseenesest täiesti konkreetne - selle raames on vähemalt kaks eeldust, mis selgitavad samasooliste seksuaalsete eelistuste „sünnipärase olemuse“ mehhanismi: (A) homoseksuaalse külgetõmbe põhjustab „eriline geen“ või geneetiline mutatsioon, teisisõnu on homoseksuaalsus kodeeritud inimese DNA-s ja edastatakse põlvest põlve; (B) homoseksuaalse külgetõmbe põhjustavad raseduse ajal esinevad kõrvalekalded (hormonaalsed või immuunsed), mis väidetavalt mõjutavad loote emakas ja põhjustavad homoseksuaalseid eelistusi beebis.
Seega jaguneb bioloogilise determinismi hüpoteesi arutelu kolmeks osaks. Esimeses osas uuritakse kriitiliselt argumente homoseksuaalsuse ja geenide suhte kohta, teises osas uuritakse kriitiliselt argumente emakasisesetest hormonaalsetest häiretest tingitud homoseksuaalse külgetõmbe arengu kohta. Kolmandas osas uuritakse kriitiliselt homoseksuaalse külgetõmbe autoimmuunse geneesi teooriat.

Esimene osa: gei geenid?
Väide homoseksuaalsuse geneetilise olemuse kohta põhineb mõnede andmete valikulisel esitamisel ja muude andmete mahasurumisel enamuse inimeste keskel, kellel puuduvad spetsiaalsed teadmised geneetika kohta. Teadus ei tunne "homoseksuaalsuse geeni", seda pole kunagi kuskil tuvastatud, ehkki katseid on olnud palju.
Mõelge uuringutele, mille põhjal LGBT + aktivistid selle argumendi esitasid. Kõigepealt tasub lühidalt kirjeldada, milliste põhimeetoditega saavad teadlased kindlaks teha, kas inimese omadus (omadus) on geneetiliselt määratud. Need meetodid hõlmavad kaksikute uurimist ja molekulaarset geneetilist analüüsi.
Kaksikute õpingud
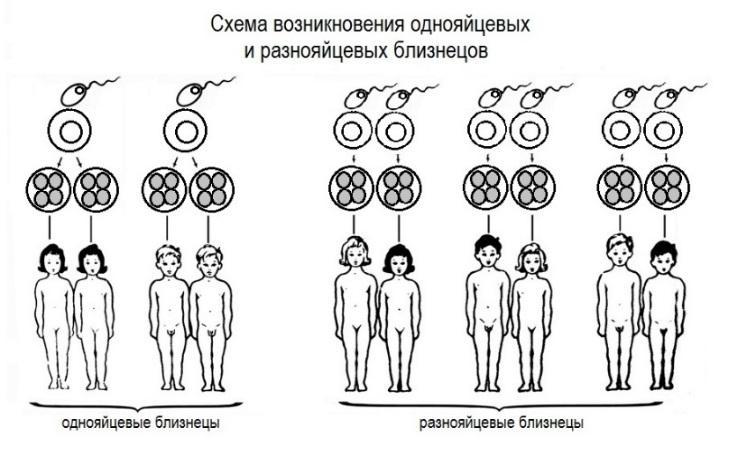
Identsete kaksikute uurimine on piisav uurimismeetod, et hinnata, kas mingitel tunnustel on geneetiline alus. Alustuseks - mida tähendab mõiste “identsed kaksikud”? Sellised kaksikud arenevad samast viljastatud munarakust, mis jaguneb osadeks, millest arenevad eraldi organismid, mis on üksteise geneetilised koopiad. Nende geenid langevad 100% -l kokku, võite neid nimetada looduslikeks kloonideks. Identseid kaksikuid nimetatakse ka identseteks või monosügootseteks (homosügootseteks) kaksikuteks. Homoseksuaalsed kaksikud moodustuvad erinevatest munadest, neid viljastavad erinevad seemnerakud. Nende geenid langevad keskmiselt kokku 50%, võib olla erinev sugu, pikkus, silmade värv, juuksed jne. Mitteidentseid kaksikuid nimetatakse ka mitteidentiteetseteks või dizügootseteks (heterosügootseteks) või kahekordseteks kaksikuteks.
Kaksikute uurimisel uuritakse kooskõla (juhus). Tunnuse kooskõla on tunnuse avaldumise tõenäosus, mis mõlemal kaksikul on. Kui identsete kaksikute mis tahes tunnus on kõrge, siis võime järeldada, et see omadus on tõenäoliselt tingitud geneetilistest teguritest. Kui tunnuse kooskõla identsetes kaksikutes ei ületa identsuste kaksikute kaksikute vastavust, näitab see, et selle tunnuse kujunemisel võib üldine keskkond olla olulisem tegur kui tavalised geenid (Yarygin 2003).
Vaja on täpselt selgitada, mida kooskõla näitab. See ei viita mingil juhul ühegi geeni olemasolule. Kaksikute tunnuse ühilduvus näitab selle tunnuse pärimisastet. Siinkohal tasub kaksikute uurimisel vaevata sõna “pärilikkus” tähenduses. Pärimine on mõõt, mis näitab, kui palju on konkreetse tunnuse varieeruvus populatsioonis (see tähendab, kui erinev see tunnus võib olla indiviiditi erinev) geenide varieeruvusega antud populatsioonis. Kaksik uuringutes ei ole pärilikkus siiski tunnuse geneetilise determinismi mõõt.
Peaaegu täielikult geneetiliselt määratud omadustel võivad olla väga madalad pärilikkuse väärtused, samas kui tunnustel, millel puudub praktiliselt geneetiline alus, võivad olla kõrged pärilikkuse väärtused. Näiteks on sõrmede arv - viis mõlemal jäsemel - inimestel geneetiliselt peaaegu täielikult kindlaks määratud. Kuid inimese sõrmede arvu iseloomustab väike varieeruvus ja täheldatud varieeruvus on enamikul juhtudel seletatav mittegeneetiliste teguritega, näiteks õnnetustega, mille tulemuseks on tunnuse vähene pärilikkuse koefitsient. See tähendab, et kui leiate kolmkümmend paari kaksikuid, kus ühel neist ei ole kätt viiel sõrmel, siis täheldatakse teise venna sama palju sõrmi äärmiselt väikeses paarisarvul, kui neid on.
Vastupidiselt võivad mõned kultuurilised jooned olla väga pärilikud. Näiteks kui kaaluda Ameerikas kõrvarõngaste kandmist kahekümnenda sajandi keskpaigas, näeksime, et seda iseloomustab kõrge pärilikkus, kuna sellest ajast sõltus see suuresti soost, mida omakorda seostatakse XX või XY kromosoomi paaride olemasoluga, seega kõrvarõngaste kandmise varieeruvus on tugevalt seotud geneetiliste erinevustega, hoolimata asjaolust, et see on pigem kultuuriline kui bioloogiline nähtus. Näiteks kui uurida kolmkümmend paari kaksikud tüdrukuid, kus üks õdedest kannab kõrvarõngaid, siis 100% juhtudest kannaks teine ka kõrvarõngaid. Täna oleks kõrvarõngaste kandmise pärandustegur madalam kui Ameerikas kahekümnenda sajandi keskpaigas mitte sellepärast, et ameeriklaste geenivaramus on toimunud muutusi, vaid seetõttu, et kõrvarõngaid kandvate meeste arv on suurenenud (Blokeeri xnumx).
Üks käitumisgeneetika pioneere oli saksa päritolu ameerika psühhiaater Franz Joseph Kallmann. Ajakirjas 1952 avaldatud artiklis ütles ta, et tema uuritud 37-i identsete (monosügootsete) kaksikute paaride korral, kui üks kaksikutest oli homoseksuaalne, siis teine oli ka homoseksuaalne, see tähendab, et kooskõlaaste oli vapustav 100% (Kallmann xnumx) Kallmann ei osutanud täpselt, kuidas ta oma uuringus osalenute monosügootsust testis. Samuti ei osutanud autor täpselt, kuidas ta uuringusse osalejaid värbas, samas kui väljaanne nentis: „potentsiaalsete osalejate otsimist korraldati mitte ainult psühhiaatriliste, paranduslike ja heategevusorganisatsioonide abiga, vaid ka otseste kontaktide kaudu põrandaaluse homoseksuaalse maailmaga” (Kallmann xnumx) Seetõttu kritiseeriti Kallmanni uuringut teravalt (Taylor 1992): Rosenthal tõi välja psühhiaatriliste probleemidega inimeste domineerimise Kallmanni vastajate hulgas (Rosenthal xnumx), Märkis Likken, et monosügootsete kaksikute ebaproportsionaalne ülekaal Callamanne'i valimis võrreldes üldpopulatsiooniga: (Lykken 1987).

Professor Edward Stein jõudis järeldusele, et Kallmanni valim "ei esindanud mingil moel homoseksuaalset elanikkonda" (Stein xnumx) Veelgi enam, Kallmann ise tunnistas, et peab oma tulemusi mitte midagi muud kui “statistilist artefakti” (Rainer 1960) Statistikas nimetatakse proove nagu Kallmanni uuringus kasutatud proove “mugavateks proovideks” - need hõlmavad objektide valimist vastavalt uurijale mugavatele kriteeriumitele. Sellist valimit kasutades ei saa teaduslikult üldistada, kuna sellise valimi omadused ei kajasta üldpopulatsiooni omadusi.
Näiteks kui küsitlus viiakse kaubanduskeskuses läbi varahommikul ainult ühe päeva jooksul, siis ei kajasta selle tulemused teiste ühiskonnaliikmete arvamust, nagu see oleks siis, kui uuring korraldataks erinevatel kellaaegadel ja mitu korda nädalas. Või kui küsite kaupluse klientidelt, kas nad alkoholi ostavad, siis reede õhtul ei lange tulemus pühapäeva tulemustega kokku.
1968-is uurisid Ameerika teadlased Heston ja Shields homoseksuaalsuse kooskõla 7-i identsetes kaksikpaarides. Uuringus osalejad leiti Madsley kaksikregistrist (Heston xnumx) Kõik vastajad olid psühhiaatrilised patsiendid. Autorid näitasid 43% -is identsuste kaksikute kooskõla. Seda uurimust kritiseeriti ka autorite endi poolt osalejate psühhiaatriliste haiguste ja äärmiselt väikese valimi tõttu (Taylor 1992; Heston xnumx).
Bailey ja Pillardi uurimus
Järgmise kaksikute seksuaalse külgetõmbe uuringu viisid 1991-is läbi Michael Bailey Northwesterni ülikoolist ja Richard Pillard Bostoni ülikoolist Ameerikas (Bailey 1991) Nad uurisid homoseksuaalsuse kooskõla erineva suguluse vendadega. Uuriti 56 paare identseid kaksikuid, 54 paare identseid kaksikuid, 142 õdesid-vendi ja vendi 57 paare2. Allolevas tabelis on toodud nende analüüsi tulemused.
Homoseksuaalne kooskõla
sõltuvalt suhte astmest (Bailey 1991)
| Suhte tüüp | Geenide protsent | Kooskõla |
| Identsed kaksikud | 100% | 52% |
| Mitte-identsed kaksikud | 50% | 22% |
| Kaksikvennad | 50% | 9,2% |
| Võsavennad (mitte sugulased) | Puuduvad olulised sarnasused | 11% |
Bailey ja Pillard tõdesid, et kuna 52% juhtudest olid identse kaksikute paari teisel õel-vennal ka homoseksuaalsed eelistused, siis "... homoseksuaalsed kalduvused tulenevad geneetilisest mõjust ...".
Nagu ka varasemates kaksikute uuringutes, on Bailey ja Pillardi uuringus põhimõttelisi probleeme. Esiteks, kui homoseksuaalsus oleks geneetiliselt määratud, oleks identsete kaksikute vastavus 100%, mitte 52%, kuna nende geenid on identsed 100%, mitte 52%. Bailey ja Pylardi artikli kommentaaris märkis Riesch ka, et geneetiliselt võõraste inimeste - poolvendade - kokkusattumus oli isegi kõrgem kui bioloogiliste kaksikvendade seas, mis näitab keskkonnamõjude olulisust. (Risch 1993) Geneetika põhimõtete kohaselt peaks lisaks 100% seksuaalsoovi kokkulangemisele identsetes kaksikutes identsete kaksikute ja mitte-kaksikute vendade kokkusattumuse protsent olema vastavalt vastavalt 22% ja 9,2% (vt tabelit allpool).
Lisaks erineb identsete kaksikute identiteet (100% geneetilisest sarnasusest) vastandlike kaksikute identiteedist (50% geneetilisest sarnasusest) 2.36 korda, kuid kui võrrelda identsete kaksikute identiteeti kaksikvendade vastavusega (50%) erinevus on järgmine: 2.39 korda, mis jällegi viitab keskkonna ilmsemale mõjule kui geneetika (vt allpool olevat tabelit).
Kategooriate vastavuse võrdlus (Bailey 1991)
| Võrdle kategooriaid | Geneetilise sarnasuse erinevus | Erinevus konkordantside vahel |
| Identsed kaksikud ja vastas kaksikud | Kaks korda nii palju tavalisi geene |
2.36
|
| Kaksikvennad ja kaksikvennad | Geenide protsendimääras pole erinevust |
2.39
|
Teiseks ei valinud Bailey ja Pillard homoseksuaalide suvalist valimit. See tähendab, et nad ei kaasanud uuringusse erapooletu akadeemilise uurimistöö standardite järgi inimesi: pole tulemustest huvitatud, ei tunne üksteist jne. Nagu teadlane parun kirjutab:
“... Selle asemel värvati osalejad geiajakirjadesse kuulutusi postitades. Selline osalejate valik on väga kaheldav, sest see sõltub selliste ajakirjade lugejaskonnast ja osalemiseks nõusolekute motivatsioonist. Selline fakt põhjustab tulemuste moonutamist, näiteks asjaolu, et homoseksuaalsete kaksikute arv on ülehinnatud. Miks? Sest osalejad võtsid enne kaksikvendade seksuaalset käitumist arvesse enne, kui nad olid nõus osalema. Ja see seab kahtluse alla valimi juhuslikkuse. Teaduslike tõendite saamiseks peaks valim olema võimalikult juhuslik, see tähendab, et uuringusse oli vaja kaasata kõik kaksikud ja seejärel viia läbi seksuaalkäitumise analüüs ... ”(Baron 1993).
Kolmandaks, nagu kirjutavad teadlased Hubbard ja Wald oma analüüsis:
"... asjaolu, et kaksikute vendade - 22% - kooskõla kahekordistab samaväärsust lihtsate vendade - 9,2% - vahel, näitab, et homoseksuaalsuse arengu põhjus pole mitte geneetika, vaid keskkond. Heterogeensete kaksikute geneetiline sarnasus on tõepoolest tavaliste vendade sarnasus. Ja kui keskkonnateguritel ja kasvatusel on heterogeensete kaksikute puhul nii suur mõju, pole üllatav, et identsete kaksikute seas on keskkonna mõju veelgi suurem. Lõppude lõpuks on identse kaksikvennaga inimese psühholoogiline ettekujutus selle kaksikuga lahutamatult seotud ... ”(Hubbard xnumx).
Teadlased Billings ja Beckwiers kirjutasid oma ülevaates "... kuigi autorid tõlgendasid järeldusi kui tõendeid homoseksuaalsuse geneetilisest alusest, usume, et tulemused näitavad vastupidi, et kasvatus- ja keskkonnategurid mõjutavad homoseksuaalsuse arengut" (Arveldused xnumx, lk 60).
Kas Bailey ja Pillardi tulemusi on korratud?
Kas kellelgi õnnestus korrata (korrata) Bailey ja Pillardi tulemusi - leida identsuste kaksikute vahel vähemalt 52% -l vastavus? 2000-is üritas Michael Bailey ise oma uurimist korrata Austraalias suures kaksikute rühmas. Homoseksuaalsete kalduvuste vastavus oli isegi väiksem kui tema esimeses uuringus. Identsete kaksikute seas oli see 20% meeste ja 24% naiste seas ning identsete kaksikute hulgas - 0% meeste ja 10% naiste3 (Bailey 2000).

Allikas: Sally Ryan ajalehele The New York Times
Rootsi epidemioloog Langström viis 2010-is läbi kaksikute seksuaalse sättumuse keeruka suuremahulise uuringu, analüüsides mitme tuhande samasooliste identsete ja heteroseksuaalsete kaksikute paari andmeid (Långström 2010) Teadlased on tuvastanud homoseksuaalseid kalduvusi samasooliste seksuaalpartnerite olemasolu suhtes kogu elu. Nad arvutasid kooskõla kahe parameetri järgi: vähemalt ühe homoseksuaalse partneri olemasolu juuresolekul elu jooksul ja homoseksuaalsete partnerite koguarvu järgi elu jooksul. Vastavuse näitajad valimis olid madalamad kui mõlemas uuringus Bailey jt. (1991) ja (2000) Osalejate rühmas, kellel oli vähemalt üks samast soost partner, oli meeste vastavuses 18% identsete ja 11% identsete kaksikute puhul; naistel vastavalt 22% ja 17%.

Allikas: Karolinska instituut
Seksuaalpartnerite koguarvust moodustasid meeste vastavuse näitajad 5% identsete ja 0% identsete kaksikute puhul; naistel vastavalt 11% ja 7%. Meestel on 61% ja 66% dispersioon seletatav keskkonnateguritega, mis mõjutavad vastavalt ainult ühte paari kaksikut, samal ajal kui dispersiooni ei selgita üldse kaksikute ühised keskkonnategurid. Unikaalsed keskkonnategurid moodustasid vastavalt 64% ja 66% hajutatuse, samas kui üldised keskkonnategurid moodustasid vastavalt 17% ja 16% (Långström 2010).
2002-is viisid Columbia ülikooli teadlased Peter Birmen ja Ameerika Yale'i ülikooli Hannah Bruckner läbi ulatusliku ja esindusliku uuringu, kus osales suur arv osalejaid (Bearman 2002).

Allikas: hannahbrueckner.com
Nad saavutasid homoseksuaalsete kalduvuste veelgi ebaolulisema taseme: 6,7% identsete kaksikute paaridena, 7,2% erinevatel identsetel kaksikutel ja 5,5% tavalistel vendadel. Birmen ja Bruckner järeldasid, et nad leiti:
"... kui on olemas olulisi tõendeid sotsialiseerumismudeli kohta indiviidi tasandil ..., näitavad meie tulemused, et laste kasvatamine soolise neutraalsuse põhimõttel, ilma et oleks selgelt kindlaks määratud lapse sugu, mõjutab homoseksuaalsete kalduvuste teket ..." (Bearman 2002).
Erinevalt äsja vaadatud töödest viisid psühhiaater Kenneth Kendler ja tema kolleegid läbi suure kaksikute uuringu, kasutades tõenäosuslikku valimit, mis koosnes 794 kaksikute paarist ja 1380 tavalistest vendadest ja õdedest (Kendler xnumx) Autorid järeldasid, et nende leiud "viitavad sellele, et geneetilised tegurid võivad tugevalt mõjutada seksuaalset sättumust". Uuring ei olnud aga piisavalt tõsine järelduste tegemiseks geenide mõju kohta seksuaalsusele: identsete kaksikute 19 paaride 324-is tuvastati homoseksuaalsete kalduvustega isik, samas kui 6 paaride 19-is olid homoseksuaalsed kalduvused ühesugused (täheldatud teine vend); samasooliste kaksikute 15-i paaridest leiti 240-is vähemalt üks homoseksuaalse kalduvusega isik, samas kui 2-i paaride 15 oli sama. Fakt, et ainult 8 kaksikpaarides 564 langesid homoseksuaalsed kalded kokku (1,4%), piirab nende tulemuste kasutamise võimalust identsete ja mitteidentsete kaksikute tõsiseks võrdlemiseks.
Tuleb meeles pidada, et identseid kaksikuid ümbritseb peaaegu sama keskkond - varajane kiindumus, suhted teiste lastega jne. - võrreldes mitteidentsete kaksikute ja tavaliste vendade ja õdedega. Kuna identsed kaksikud on välimuse ja iseloomu poolest sarnased, on sama suhtumine neisse tõenäolisem kui identsete kaksikute ja tavaliste vendade ja õdede suhtes. Seetõttu on mõnel juhul kõrgemat kooskõlalisuse koefitsienti seletatav pigem keskkonna kui geneetiliste teguritega.

Professor Kenneth Kendler.
Allikas: Virginia Rahvaste Ühenduse ülikool.
Psühhiaater Jeffrey Satinoveri (Satinover xnumx) tegurid, mis mõjutavad täielikult inimese seksuaalkäitumistüübi kujunemist, võib jagada viide kategooriasse:
1) emakasisene (sünnieelne) toime, näiteks hormoonide kontsentratsioon;
2) emakaväline (sünnitusjärgne) füüsiline mõju, näiteks trauma ja viirusnakkused;
3) kehavälised kogemused, näiteks perekondlikud suhted, haridus;
4) sünnieelne kogemus, näiteks stereotüüpse korduva käitumise tugevdav toime;
5) valik.

Allikas: ihrc.ch
100% -lise vastavuse puudumine identsetes identsetes kaksikutes viitab mitte ainult sellele, et geneetiliste tegurite mõju on tühine, vaid ka sellele, et mittegeneetilised tegurid ei saa olla eranditult emakasisesed. Lõppude lõpuks, kui see oleks nii, siis oleks kooskõla ikkagi 100% lähedal, kuna identseid kaksikuid mõjutavad emakasisese keskkonna samad tegurid ”(Satinover xnumx, lk 97).
Kui geenid mängivad rolli inimeste eelsoodumuse kujundamisel teatud seksuaalsete soovide ja käitumise suhtes, siis võimaldavad kõik need uuringud kindlalt väita, et geneetiliste tegurite mõju ei ammenda seda teemat. Kaksikute uurimistööd kokku võttes võime julgelt järeldada, et teadus ei ole tõestanud, et seksuaalne iha üldiselt ja eriti homoseksuaalsed kalduvused on määratud inimese geenide poolt.
Molekulaargeneetilised uuringud
Uurides geneetika osalemist homoseksuaalsete kalduvuste kujunemises ja võimaluse korral selle osaluse astet, oleme seni uurinud uuringuid, milles tunnuse geneetiline pärimine (konkreetsel juhul homoseksuaalse külgetõmbe korral) on määratud klassikalise geneetika abil, kuid nad ei seadnud ülesandeks kindlaks teha, milline selle tunnuse eest vastutavad spetsiifilised geenid. Samal ajal saab geneetikat uurida nn. molekulaarsed meetodid, mis võimaldavad kindlaks teha, millised konkreetsed geneetilised variandid on seotud füüsiliste või käitumuslike tunnustega.
Dean Haymeri uuring
Dean Haymer ja tema kolleegid Ameerikas Marylandis asuvas Riiklikus Tervise Instituudis tegid ühe esimestest katsetest läbi viia homoseksuaalsete kalduvuste molekulaargeneetiline analüüs (Hamer 1993) Haymer uuris peresid, kus olid identsed isased kaksikud, kus vähemalt ühel kaksikutest olid samasoolised külgetõmbejõud. Perekondade koguarvu hulgast tuvastas Haymer 40i, kus homoseksuaalsel vennal oli erinev vend, kes oli samuti homoseksuaalne, ja uuris nende DNA-d sarnaste saitide osas. Sarnast uuringut nimetatakse “seotud pärimise uurimiseks” - inglise keeles “genet linkage study”.
Lingitud pärimise uurimisel tehakse järgmist: rühmas katsealustel, kellel on ühine teadaolev omadus, viiakse läbi analüüs sarnaste DNA sektsioonide olemasolu kohta - neid nimetatakse markeriteks. Kui selgub, et katsealuste rühmas asub suur arv markereid samas DNA piirkonnas, siis võib eeldada, et kõik need markerid on päritud “koos” - seotud - st nad võivad olla osa mingist geenist (Pulst 1999).
Haymer ütles, et 33-ist pärinevates paarides 40 on homoseksuaalsetel vendadel X-kromosoomis sama soo piirkond, mida ta nimetas "Xq28". Heimer järeldas, et Xq28 piirkond sisaldab geene homoseksuaalsete kalduvuste jaoks.

vastuoluliste artiklite autorid -
geneetikat ja seksuaalsust käsitleval konverentsil
Mai 1995 (Soomlane 1996)
Esiteks tuleb märkida, et Haymeri tulemusi tõlgendatakse sageli valesti. Paljud inimesed arvavad, et Haymer leidis kõigist 28-i paaridest ja kõigist 33-i meestest identse DNA piirkonna - Xq66 -, kuid tegelikult leiti, et Xq28 piirkonna nukleotiidijärjestused olid igas kaksikvennas vendade vahel identsed ja Xq28 järjestus kõigis paarides polnud identne - Haymer ei leidnud kurikuulsat gei-geeni.
Sellel uuringul on mitmeid olulisi puudusi. Haymer ei kontrollinud Xq28 kokkusattumust heteroseksuaalse külgetõmbega paarides, vaid ainult homoseksuaalide seas (Byne xnumx) Kui ta ei leia seda saiti heteroseksuaalsete vendade hulgast, vaid ainult homoseksuaalide seas, räägiks see tema järeldust toetavast tulemusest. Kui ta oleks aga oma heteroseksuaalsete vendade hulgast avastanud Xq28, oleksid tema järeldused omandanud nullväärtuse (Horton xnumx) Samuti, nagu teadlased Fausto-Sterling ja Balaban märkisid, sisaldab Heimeri proov mittetäielikku hulka andmeid: 40 juhtudest mõõdeti otseselt ainult 15 DNA heterosügootsuse karakteristikuid; ülejäänud 25 juhtumite korral arvutati andmed kaudselt (Fausto-Sterling 1993) Ainult 38% juhtudest mõõtsid Heimer jt otseselt ema X-kromosoomi heterosügootsuse taset ja 62% -l arvutasid nad selle lihtsalt olemasolevate andmebaaside põhjal.
Tuleks mainida järgmist episoodi, mis on seotud aasta Haymer 1993 avaldamisega. New York Native ajakiri avaldas 1995-is artikli pealkirjaga “Homosuhtluse“ geenide uurimine ”ei pidanud proovile: Chicago Tribune ajakirjanik John Krudson paljastas uurija poolt võimaliku teadusliku võltsimise” (Chicago Tribune 1995) Artiklis märgitakse, et erinevad teadlased kritiseerisid Haymeri tööd tõsiselt selle pärast, et Haymer ei kontrollinud Xq28 olemasolu heteroseksuaalsete vendade seas. Kriitikute hulka kuulusid tuntud bioloogid ja geneetikud Richard Levontin ja Ruth Hubbard Harvardi ülikoolist (Chicago Tribune 1995) Veelgi enam, samas artiklis öeldakse, et Tervise Instituudi föderaalne eetikabüroo uurib ühe Heimeri labori noore töötaja kaebust, kelle nime ei teata, kes teatas Heimeri uuringus tehtud tulemuste taglasest: selle ohvitseri ütluse kohaselt oli Heimer teadlikult jättis publikatsioonist välja tulemused, mis osutavad homoseksuaalsete kalduvuste geneetilise ettemääratluse teooria põhjendamatusele (Chicago Tribune 1995) Mõni kuu pärast artikli avaldamist New Yorgi põliselanikus avaldas ajakiri Scientific American veel ühe artikli, milles kinnitati föderaalse eetikabüroo Heimeri (Horgan xnumx, lk 26). Riiklik terviseinstituut ei avaldanud uurimise tulemusi, kuid Haymer viidi hiljem teise osakonda. Samuti tuleb märkida, et Haymer viis läbi uuringu „homoseksuaalsuse geeni” kohta, kasutades selleks stipendiumi, mis eraldati tegelikult Kaposi sarkoomi, nahavähi, mis mõjutab sageli AIDSi homoseksuaalseid patsiente (Mukherjee xnumx, lk 375). Haymeri väljaande kehtivus sõltus sellest, kas sõltumatu teadlaste meeskond võiks saada samu tulemusi. Seda ei juhtunud.

Haymeri tulemuste korratavus
1999-is viis Lääne-Ontario ülikooli teadlaste rühm, keda juhtis teadlane Rice, läbi sarnase uuringu (kasutades “geneetilise sideme meetodit”) 52-i geimehi (Riis xnumx) Autorid ei suutnud Haymeri saadud tulemusi korrata ja järeldasid: "meie uuringu tulemused ei paljastanud mingeid tõendeid meeste homoseksuaalsuse ja geenide vahelise seose kohta".
Seejärel viidi 2005-is läbi uus uuring Dean Haymeriga (Mustanski Xnumx) Autorid ei leidnud statistiliselt olulist seost Xq28i ja homoseksuaalsete kalduvuste vahel, kuid väitsid, et nad leidsid teiste saitide (7, 8 ja 10 kromosoomides) huvitava korrelatsiooni.
Neid tulemusi ei saanud aga korrata ühes teises 2009i uuringus, kui Inglismaa Oxfordi ja Kanada Ontario ülikooli teadlaste rühm viis läbi uuringu 55 perekondade kohta, milles olid geid: 112 osalejatelt koguti geneetilist materjali ja viidi läbi genoomi hõlmav seoste otsimine koos 6000 geenimarkerite kaasamisega (Ramagopalan 2010) Analüüs ei tuvastanud statistiliselt olulist seost geneetiliste markerite ja homoseksuaalsuse vahel.
2015-is väitis Ameerika erinevate teaduskeskuste autorite rühm genoomi hõlmavate assotsiatsioonide otsingu järgi, et nad leidsid 8-i kromosoomis oleva saidi jaoks olulise seose ja Xq28-i jaoks vähem olulised (Sanders xnumx) Oma artikli järeldustes tunnistasid autorid, et "geneetiline mõju homoseksuaalsetele kalduvustele pole kaugeltki määrav ... tõenäoliselt on see mõju osa mitmefaktorilisest põhjusest".
2017-is rakendas sama autorirühm moodsamat ja täpset meetodit, mida nimetatakse genoomi hõlmavaks seoste otsimiseks4. Geenivaramu assotsiatsioonide otsimine põhineb genoomi järjestamise tehnoloogia kasutamisel (DNA-st teabe lugemine), et teha kindlaks DNA spetsiifilised tunnused, mis võivad olla seotud uuritava tunnusega. Teadlased uurivad miljonite geneetiliste variantide arvukust paljudel ühise tunnusega isikutel ja isikutel, kellel seda omadust pole, ning võrdlevad mõlema rühma geneetiliste variantide sagedust. Eeldatakse, et need geneetilised variandid, mis on tunnuse omanike seas tavalisemad kui ilma selleta, on kuidagi selle tunnusega seotud. Seekord leiti statistiliselt olulised seosed 13 ja 14 kromosoomi piirkondade (Sanders xnumx).

Sandersi ja tema kolleegide (2017) uuringus ei leitud homoseksuaalsete kalduvuste geeni ega tõestatud nende geneetilist seisundit (autorid ise lükkavad selle ümber) ega kinnitanud ka aasta Haymer 1993 tulemusi, mis pani aluse pikale homoseksuaalsuse geenidele. Selle väljaande üks järeldusi oli eeldus, et kõik ülalnimetatud geneetilised variandid võivad mõjutada eelsoodumus homoseksuaalsed kalduvused (Sanders xnumx, lk 3).
Inimgenoomi dekodeerimise projektijuht Francis Collins kirjutab järgmist:
„Tõenäosus 20% lähedal, et ka homoseksuaalse mehe identne kaksik on homoseksuaalne (võrreldes 2 - 4% -ga üldpopulatsioonis) näitab, et seksuaalset sättumust mõjutavad geenid, kuid nad pole DNAsse sisse lülitatud, ja kõik seotud geenid esindavad eelsoodumust, kuid mitte eeldatavat järeldust ... ”(Collins 2006).
Ameerika inimgeneetika seltsi 2012-i aastakonverentsil esitleti eriti suurt uuringut genoomi hõlmavate ühenduste otsimise meetodi kohta, mille eesmärk oli kindlaks teha homoseksuaalsete kalduvustega seotud geneetilised variandid.Drabant 2012) Kogu genoomi hõlmava otsingu tulemusel ei leitud mõlemast soost olulisi suhteid homoseksuaalsete kalduvuste osas. Samal ajal uuriti ettevõtte andmebaasist 23andMe tuhandeid isikuid.
Kõige uuemate ja suurimate autorid teadustöö homoseksuaalsuse geneetikast rääkinud selle tulemuste kohta:
"Inimese seksuaalkäitumist on genoomi põhjal praktiliselt võimatu ennustada,"
ütleb uuringu kallal töötanud Massachusettsi üldhaigla analüütilise ja translatiivse geneetika osakonna professor Ben Neal.
California ülikooli geneetikainstituudi professori David Curtise sõnul
"Inimese populatsioonis ei ole ühtegi geenikombinatsiooni, millel oleks oluline mõju seksuaalsele orientatsioonile. Inimese seksuaalkäitumist on genoomi põhjal praktiliselt võimatu ennustada."
Epigeneetika
Los Angelese California ülikooli teadlaste rühm 2015 tutvustas Ameerika inimgeneetika seltsi konverentsil kokkuvõtet5mis väitis, et teadlased suutsid epigeneetiliste markerite põhjal tuvastada seksuaalse eelistuse täpsusega 67% (Ngun jt. 2015). Oma tööle maksimaalse tähelepanu äratamiseks korraldasid autorid isegi ajakirjandust hõlmava pressiteate (ASHG 2015) Vaatamata uuringu avatud vastuolulisele olemusele ja kahtlasele vahendamismeetodile levisid uudised kohe peavoolu ajalehtede pealkirjadesse (Yong xnumx).
Epigeneetika on teadus, mis uurib nähtusi, kus geenide ekspressioon muutub tänu mehhanismidele, mis ei mõjuta geenides DNA järjestuse muutumist. Teisisõnu, epigeneetilised protsessid on protsessid, milles geeniekspressiooni astet (see tähendab keha füsioloogilisi omadusi) mõjutavad muud tegurid. DNA molekuli ruumiline konfiguratsioon võib mõjutada geeni ekspressiooni (ekspressiooni) ja selle konfiguratsiooni määravad spetsiaalsed regulatoorsed valgud, DNA-ga seotud ensüümid. Üks mõjumehhanisme on DNA metüleerimine. Regulatiivsete valkude ja DNA kombinatsiooni nimetatakse epigeneetiliseks markeriks.
Noored ja tema kolleegid väitsid, et nende uuringu peamine eesmärk oli testida võimalust epigeneetiliste markerite abil indiviidi seksuaalset sättumust kindlaks teha. Sel eesmärgil uurisid nad identsete kaksikvendade 37 paari DNA proove, kõigis neis paarides oli üks vend homoseksuaalne ja 10 paari identseid kaksikvenda, kusjuures mõlemad vennad olid homoseksuaalsed. Nagu kokkuvõttes öeldud, uurisid teadlased arvutistatistika algoritmi FuzzyForest kasutades mitmesuguseid klassifitseerimismudeleid (heteroseksuaalsed vs homoseksuaalsed) ja valisid lõpuks kõige paremini toimiva mudeli, sealhulgas 5 epigeneetilised markerid, mis klassifitseerisid objektid 67% -l juhtudest. Autorid soovitasid seksuaalseid eelistusi kontrollida 5 epigeneetiliste markerite abil. Kuid selline tõlgendus põhjustas pehmelt öeldes ekspertide kriitika puhkemise (Teadusmeedia keskus 2015, Roheliselt xnumx, Yong xnumx, Gelman 2015, Briggs 2015) Metoodika (äärmiselt madal valimivõimsus, kahtlane statistiline lähenemisviis suure valede positiivsete tulemuste riskiga jne) ja selle tõlgendamine tekitas suuri kahtlusi. John Grillie Albert Einsteini meditsiinikolledži epigenoomikakeskusest märkis, kommenteerides Nguni ja tema kolleegide uuringu ümbritsevat hüpet:
„... rääkimata temast või tema kolleegidest isiklikult, kuid kui tahame seda teadusvaldkonda säilitada, ei saa me enam lubada halbade epigeneetiliste uuringute usaldamist. "Halva" all mõtlen ma tõlgendamatut. ... "(Roheliselt xnumx).

Lõpuks seati isegi kahtluse alla retsensentide objektiivsus, kes jätsid selle jätkamise konverentsil esitlemiseks, ja artiklit muidugi ei avaldatud kunagi kuskil.
Miks on molekulaargeneetiliste uuringute tulemused nii vastuolulised - muutuvad ja varieeruvad?
Geneetika piiratud roll
Tõendeid homoseksuaalsete kalduvuste geneetilise olemuse kohta ei saa kinnitada. Teadus ei tunne "homoseksuaalsuse geeni". Selle sajandi alguses käivitati suuremahuline rahvusvaheline projekt “Inimese genoomi projekt” - projekt “Inimese genoom”. Selle raames viidi läbi inimese geneetiliste kaartide koostamine - milline geen, millises kromosoomis asub, milliseid valke see kodeerib jne. Igaüks saab kontrollida - homoseksuaalsuse geene pole seal näidatud (Inimese genoomi ressursid NCBI-s).
Mayer ja McHugh kirjutavad oma töös järgmist:
“... Nagu inimese käitumisomadustega seoses on juba mitu korda kinnitatud, on geneetilise teguri mõju homoseksuaalse kalduvuse või käitumisharjumuste kalduvusele võimalik. Geenide fenotüübiline avaldumine sõltub tavaliselt keskkonnateguritest - erinev keskkond põhjustab erinevate fenotüüpide moodustumist isegi samade geenide jaoks. Seetõttu, isegi kui mõned geneetilised tegurid mõjutavad homoseksuaalseid kalduvusi, mõjutavad seksuaalseid eelistusi ja kalduvusi ka mitmed keskkonnategurid, sealhulgas sellised sotsiaalsed stressifaktorid nagu psühholoogiline ja füüsiline vägivald ning seksuaalne ahistamine. Seksuaalsete huvide, soovide ja ajendite kujunemisest terviklikuma pildi saamiseks tuleb arvestada arengu, keskkonna, kogemuste, ühiskonna ja tahte teguritega. (Näiteks on sotsiaalgeneetikud registreerinud geenide kaudse rolli eakaaslastega käitumises, mis näitab, et inimese välimus võib mõjutada aktsepteerimist või tagasilükkamist konkreetses sotsiaalses rühmas (Ebstein 2010).
Kaasaegne geneetika teab, et geenid mõjutavad indiviidi huvide ringi ja tema motivatsiooni ning vastavalt mõjutavad kaudselt käitumist. Ehkki geenid võivad seega inimese teatud käitumisviisidesse petta, on tema võime toiminguid otseselt juhtida, hoolimata paljudest muudest teguritest, väga, väga ebatõenäoline. Nende mõju käitumisele on peenem ja sõltub keskkonnategurite mõjust ... "(Mayer 2016).
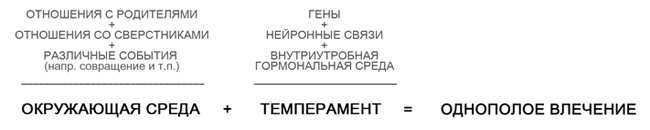
Sissesündinud tegurid, mis võivad orienteerumist mõjutada, hõlmavad temperamendiomadusi nagu kerge ja haavatav iseloom, suurenenud emotsionaalne tundlikkus, häbelikkus, passiivsus jne. Teadlased ise, mille tulemusi kasutatakse LGBT + aktivistide retoorikas - liikumised, ei julge väita, et homoseksuaalsuse määravad geenid, parimal juhul usuvad nad, et samasooliste ligitõmbamine on seotud bioloogiliste ja keskkonnategurite kombinatsiooniga, kus viimastel on suur roll . Seda, et homoseksuaalsus on kaasasündinud, kuuleme peamiselt Hollywoodi filmides, vestlussaadete reitingus, lugudes või sotsiaalvõrgustike kommentaarides. Teadusringkondades pole aga tegelikult ühtegi kohusetundlikku uurijat, kes ütleks, et on leidnud homoseksuaalse külgetõmbe geneetilise või mõne muu bioloogilise põhjuse.
Uuringud, mille eesmärk on kindlaks teha, kas geenid (eriti saidil Xq28) eksisteerivad samasooliste seksuaalse ihaga. Koostanud V. Lysov (2018)
| Allikas ja proovide võtmine |
Meetod analüüs |
Tulemused avaldamise järgi | Kas on tõendeid Xq28 markerite ja homoseksuaalsuse vahelise seose kohta? | Muud tulemused |
| Dean Hamer jt. Xnumx 40 perekonnad, millest igaüks koosnes homoseksuaalsest probandist ja tema sugulaste hulgast valitud homoseksuaalidest |
seotud pärimisuuringud | 33 perekondade 40 juhtumitel langesid X kromosoomi piirkonnas q28 asuvad geneetilised markerid kokku | tinglikultKuid kolleegid kritiseerivad meetodeid ja tõlgendamist: Baron 1993; Pool 1993; Fausto-Sterling jt. Xnumx; Terav 1993; Byne xnumx; McLeod 1994; Norton 1995, Haymerit ennast kahtlustati võltsimises: Horgan xnumx | - |
| Jennifer Macke jt. Xnumx 36 perekonnad, millest igaüks koosnes homoseksuaalsest probandist ja tema sugulastest, kelle hulgas oli vähemalt üks homoseksuaalne vend |
kandidaatgeenide otsimine - androgeeniretseptori geen (X-kromosoom) | statistiliselt olulisi seoseid valimis ei leitud | - | puudub seos androgeeni retseptori geeniga (X-kromosoom) |
| Stella Hu jt. Xnumx (teadusrühm Dean Hamer) 33 perekonnad, millest igaüks koosnes homoseksuaalsest probandist ja tema sugulastest, kelle hulgas oli vähemalt üks homoseksuaalne vend |
seotud pärimisuuringud | 22 perekondade 32 juhtumitel langesid X kromosoomi piirkonnas q28 asuvad geneetilised markerid kokku | tinglikultvaata Hamer 1993 | - |
| George Rice jt. Xnumx 46 perekonnad, millest igaüks koosnes homoseksuaalsest probandist ja tema sugulastest, kelle hulgas oli vähemalt üks homoseksuaalne vend |
seotud pärimisuuringud | X-kromosoomi q28 piirkonnas asuvad geneetilised markerid ei vastanud | ei | - |
| Michael DuPree jt. Xnumx (teadusrühm Dean Hamer) 144 perekonnad, millest igaüks koosnes homoseksuaalsest probaadist, kellel oli vähemalt üks homoseksuaalne vend |
kandidaatide geenide otsimine - aromataasi geen CYP15 (15 kromosoom) | statistiliselt olulisi seoseid valimis ei leitud | - | puudub seos aromataasi geeniga CYP15 (15-I kromosoom) |
| Mustanski jt. Xnumx (teadusrühm Dean Hamer) 146 perekonnad (sealhulgas perekonnad Hamer 1993 ja Hu 1995 uuringutest), millest igaüks koosnes homoseksuaalsest probandist, kellel oli vähemalt üks homoseksuaalne vend |
seotud pärimise genoomi hõlmav uuring | proovis leiti statistiliselt oluline seos 7 kromosoomi markeriga ja autorite sõnul 8 ja 10 kromosoomide markerite “lähedane tõenäolise olulisuse kriteeriumidele”. | ei | suhtlus 7-kromosoomi markeritega vastavalt Landeri ja Kruglyaki (1995) kriteeriumitele, mis on LOD parim indikaator* võrdsustatud xnumx |
| Sreeram Ramagopalan jt. Xnumx (George Rice'i teadusrühm) 55 perekonnad, millest igaüks koosnes homoseksuaalsest probaadist, kellel oli vähemalt üks homoseksuaalne vend |
seotud pärimise genoomi hõlmav uuring | statistiliselt olulisi seoseid valimis ei leitud | ei | Landeri ja Kruglyaki (7) kriteeriumide kohaselt 1995-i kromosoomis olevate markeritega seoseid ei leitud |
| Binbin Wang jt. Xnumx rühm Xnumx homoseksuaalseid mehi ja kontrollrühm Xnumx heteroseksuaalseid mehi |
kandidaatgeenide otsimine - heliheki (SHH) geen (7 kromosoom) | statistiliselt olulisi seoseid valimis ei leitud | - | leidsid statistiliselt olulise rühmadevahelise erinevuse rs9333613 geeni positsioonis olevate mutatsioonide suhte osas, mida autorid tõlgendasid kui "võimaliku seose olemasolu geeni mutatsioonide ja samasooliste ligitõmbe vahel" |
| Emily Drabant jt. Xnumx 7887i mehed ja 5570i naised (ei ole sugulusega seotud), kellel on Kleini küsimustiku kohaselt tuvastatud sugutung ja enesetunnetus |
täielik genoomiühenduse otsing | valimis ei leitud statistiliselt olulisi (5 × 10 - 8) seoseid | ei | statistiliselt olulisi seoseid ei leitud |
| Sanders jt. Xnumx 384 perekonnad, millest igaüks koosnes homoseksuaalsest probaadist, kellel oli vähemalt üks homoseksuaalne vend |
seotud pärimise genoomi hõlmav uuring | proovis leiti statistiliselt oluline seos markeriga 8 kromosoomis ja tõenäoline seos Xq28-iga | tinglikult: vastavalt Landeri ja Kruglyaki (1995) kriteeriumitele olid Xq28 markerite parimad LOD indikaatorid võrdsed 2,99-iga, mis vastab oletatavale väärtusele ("sugestiivne olulisus") | suhtlus 8 kromosoomi markeritega vastavalt Landeri ja Kruglyaki (1995) kriteeriumitele; parim LOD skoor oli 4,08 |
| Sanders jt. Xnumx rühm 1077 homoseksuaalseid mehi ja 1231 heteroseksuaalseid mehi (samad subjektid nagu Sanders jt. 2015) |
täielik genoomiühenduse otsing | valimis ei leitud statistiliselt olulisi (5 × 10 - 8) seoseid | ei | statistiliselt olulisi seoseid ei leitud. Autorid märkisid, et 13 ja 14 kromosoomide markerite jaoks saadi oluliseks lähenevad väärtused |
* LOD = koefitsientide mitmepunktiline logaritm, vt Nyholt DR. Kõik LOD-d pole loodud võrdselt. Am J Hum Genet. 2000 august; 67 (2): 282 - 288. http://doi.org/10.1086/303029. Statistiliselt oluline LOD geeniuuringutes on ≥3,
Nagu üks Ameerika blogijatest tabavalt ütles: "... katsed homoseksuaalsust bioloogiliselt seletada sarnanevad iPhone'idega - igal aastal ilmub uus ..." (Allen 2014) Lõpuks, ilmselt homoseksuaalsete kalduvuste edendajate seisukohalt, loosung "Tõenäoliselt sündinud eelsoodumusega"6 on hoopis teistsuguse propagandaefektiga.

Alkoholismi geeni on üritatud tuvastada (Taastumisküla 2017; NIAAA 2012) ja “tapjageen” (Davis 2016; Parshley xnumx), aga nagu “homoseksuaalsuse geeni” puhul, ei leitud tõendeid väite toetamiseks, et “sellised on sündinud”. Adekvaatsel inimesel poleks ühelt poolt mõtet alkoholismi ja mõrvu õigustada geenide mõjul - lõppude lõpuks määravad need nähtused valiku, mitte ettemääratud. Loo “pioneer” koos homoseksuaalsuse geeniga Dean Haymeril on ilmselgelt suurepärane äriline talent, kes tegutseb oskuslikult avaliku moe raames. Haymer, olles veidi aega pärast oma 1993-i artikli avaldamist, avaldab raamatu “The Passion Science: Search for Homosexuality Genes and Behevioral Biology”, mis tegi vahet LGBT + liikumise seas (Hamer 1994) ja tõi talle märkimisväärset kasumit. Kümme aastat hiljem teeb Haymer uue sensatsiooni, avaldades raamatu pealkirjaga “Jumala geen: kuidas meie geenid usu ette tellivad” (Hamer 2004), milles ta avaldas arvamust, et usklikud on peaaegu geneetilised mutandid (V.L .: on naljakas jälgida sellist selektiivsust kahe geneetilise hüpoteesi osas: homoseksuaalsete kalduvuste väidetav geneetiline tinglikkus on esitatud positiivses valguses , ning geenide ja religiooni väidetav seos on negatiivne nagu mutatsioon.). Loomulikult ei ole Heimeri hüpoteesidele tänaseks päevaks kinnitust leitud, kuid ka tema teooria võeti LGBT + kogukonnas väga soojalt vastu, Ameerika ajakiri Time avaldas selleks puhuks isegi spetsiaalse kaane.

Seejärel lahkus Dean Haymer teadusest ja keskendus ühiskondlik-poliitilisele tegevusele: koos oma "mehe" Joseph Wilsoniga (New York Times 2004) ta asutas filmistuudio "QWaves", mis on spetsialiseerunud toodetele, mis keskenduvad liikumisele "LGBT +" (Huffpost 2017).
Kuulus bioloog ja teaduse populariseerija Richard Dawkins iseloomustab filosoofiliselt homoseksuaalsuse geneetilise determinismi hüpoteesi:
„… Mõningaid keskkonna tingimusi on lihtne muuta. Teised on rasked. Mõelge, kui sügavalt oleme seotud oma lapsepõlve aktsendiga: täiskasvanud immigrant sildistatakse kogu elu välismaalasele. Siin on palju rangem determinism kui enamiku geenide tegevuses. Oleks huvitav teada statistilist tõenäosust, et laps, kes on kokku puutunud teatud keskkonnamõjuga, näiteks kloostri usuõpetusega, saab hiljem sellest mõjust vabaneda. Sama huvitav oleks teada statistilist tõenäosust, et mees, kellel on Xq28 piirkonna X-kromosoomis kindel geen, on homoseksuaalne. Lihtne demonstreerimine, et on olemas geen, mis "viib" homoseksuaalsuseni, jätab selle tõenäosuse olulisuse küsimuse peaaegu täiesti lahtiseks. Geenidel pole determinismi monopoli ... "(Dawkins xnumx, lk 104).
Vene seksoloogia üks silmapaistvamaid tegelasi, professor Georgy Stepanovich Vasilchenko, rääkides homoseksuaalsete kalduvuste tekkimise põhjustest, osutab järgmisele:
„... aju diferentseerumise häired ja hormonaalsed nihked ei määra siiski homoseksuaalse külgetõmbe tekkimist, vaid saavad aluseks seksuaalse identiteedi ja soorollikäitumise moonutustele, mis suurendavad homoseksuaalsuse riski. Neuroendokriinne varustus on ainult libiido energiakomponent. Homoseksuaalsuse teket soodustavad ka perverssusele omased etioloogilised tegurid ja patogeneetilised mehhanismid ... "(Vassilchenko 1990, lk 430).
Meeste homoseksuaalsuse geneetilise teguri hüpotees, pakkudes naistele evolutsioonilist eelist
Eraldi tasub mainida Itaalia teadlaste veidrat hüpoteesi, mis nende sõnul "Ei sobi ühegi olemasoleva homoseksuaalsuse geneetilise mudeliga". Eeldus, et homoseksuaalsuse põhjustavad geenid, on vastuolus loodusliku valiku põhimõttega, mille kohaselt järglaste tootmiseks vajalike heteroseksuaalsete funktsioonide elluviimist takistavate geenikandjate arv peaks pidevalt vähenema, kuni see täielikult kaob. Kuid nagu näidatud statistika, kasvab homoseksuaalseks pidavate inimeste arv iga põlvkonnaga. Põhjus on selge: homoseksuaalsus ei ole geneetiliselt ajendatud, kuid ei soovinud leppida ilmselge Camperio-Ciani-ga ja kolleegid tulid välja keeruka selgitusega, mis pidi hakkama saama “Darwini paradoksiga”. Nende hüpotees viitab teatud "X-kromosomaalse faktori" olemasolule, mis emaliini kaudu edasi kandudes võib suurendada mõlemast soost androfiiliat (meeste seksuaalset külgetõmmet), suurendades sellega naiste viljakust, kompenseerides meeste vähenenud viljakust (Camperio-Ciani 2004).
See hüpotees võiks väita teatavat usaldusväärsust, kui teadlased leidsid sobivad kompensatsioonitasemed - näiteks kui heteroseksuaalsete järglastega emal oli 2-laps ja homoseksuaalse järglasega emal oli 4. Tegelikult osutus erinevus väheoluliseks: keskmiselt oli lapse 2,07 esimeses ja 2,73 - teises (34% rohkem) ja seda vaatamata asjaolule, et homoseksuaalide ja heteroseksuaalide paljunemise tasemed olid peaaegu 5 korda erinevad: vastavalt 0,12 ja 0,58 (vastavalt 383) % vähem) (Iemmola xnumx). Teadlased selgitavad heteroseksuaalide ebatavaliselt madalat viljakust sellega, et kontrollrühmana pidid nad olema homoseksuaalsete probandidega võimalikult sarnased ja seetõttu olid enamik neist vallalised. Kuid isegi kui võtta need mitteametlikud andmed, selgub, et piisava hüvitise saamiseks vajavad homoseksuaalsete järglaste emad rohkem kui 7 last ... Lisaks ei leitud olulist erinevust eelmise põlvkonna (vanavanemate) viljakuses, mis ei nõustu samuti teesiga geneetilise geneetika kohta üleandmine.
Proovin andmeid selgitada, märgivad autorid, et homoseksuaalidel on kalduvus liialdada sugulaste seas mitteheteroseksuaalide arvuga, heteroseksuaalide arv aga vastupidi väheneb, mis võib tulemuste osas erineda. Nad ütlevad ka, et viljakuse erinevusi saab seletada füsioloogiliste või käitumuslike põhjustega, näiteks madalama abordimääraga või suurenenud võimega partnereid leida. Lõpuks autorid rõhutadasee suurenenud emade viljakus seletab vähem kui 21% meeste valimisse kuuluvate meeste seksuaalse sättumuse erinevustest.
„See on kooskõlas teoreetiliste ja empiiriliste uuringutega, mis näitavad, et isiklik kogemus on võimas tegur inimese seksuaalkäitumise ja enesemääratluse kindlaksmääramisel. Võimalik, et emade homoseksuaalsuse kõrgem tase tuleneb pigem kultuurilistest, mitte geneetiliselt päritud joontest. Paljudes ühiskondades, näiteks Põhja-Itaalias, veedavad emad oma lastega palju aega, eriti esimestel aastatel, mis on ülioluline seksuaalse identiteedi ja orientatsiooni kujunemiseks. See viitab sellele, et ema ja tema pere võivad olla lapse teatavate käitumisharjumuste ja hoiakute, sealhulgas tulevaste seksuaalsete eelistuste ja käitumisega seotud tunnuste peamiseks allikaks ”(Camperio-Ciani 2004).
Pärast 3-i uuringute läbiviimist olid autorid sunnitud tunnistama, et saadud andmed "Need ei võimalda meil kindlaks teha, kuivõrd oletatav X-kromosoomi tegur viib inimese homo- või biseksuaalsusele või isegi eelsoodustab seda" (Ciani xnumx) Lühidalt, nende uuringute panus homoseksuaalse külgetõmbe geneesi mõistmisse on null.
30.08.2019i autoriteetsetes teadusväljaannetes avaldatud kõigi aegade suurim geeniuuring teadus, tuvastas umbes 500 tuhande inimese valimi põhjal, et enam kui 99% homoseksuaalsest käitumisest on määratud sotsiaalsete ja keskkonnategurite poolt. Vastavalt California ülikooli geneetika instituudi professor David Curtis: "See uuring näitab selgelt, et gei geeni pole olemas." Inimeste hulgas pole sellist geenide kombinatsiooni, millel oleks oluline mõju seksuaalsele orientatsioonile. Tegelikult on inimese seksuaalset käitumist tema genoomi järgi võimatu ennustada. ”
Teine osa: hormoonid?
Lisaks geneetika mõjule osutavad liikumise “LGBT +” aktivistid väidetavalt emakasisesele kokkupuutele homoseksuaalse külgetõmbe bioloogilise geneesi väidetava mehhanismina. On arusaadav, et perioodil, kui loode on emaüsas, toimib lootele tegur (hormoonid või immuunsuse antikehad), mis häirib selle normaalset arenguprotsessi, mis viib veelgi homoseksuaalse külgetõmbe väljakujunemiseni.
Hormonaalse mõju hüpoteesi seksuaalse eelistuse kujunemisele testimiseks uurime emakasisese hormooni kontsentratsiooni kehalisele arengule ja poistele või tüdrukutele tüüpilise käitumise kujunemise seoseid varases lapseeas. Hormonaalse emakasisese tasakaalustamatuse eksperimentaalset modelleerimist muidugi eetilistel ja praktilistel põhjustel inimestel ei tehta, kuna hormonaalsed häired põhjustavad olulisi anatoomilisi ja füsioloogilisi kõrvalekaldeid, on see võimalik ainult katseloomadel7. Sellegipoolest on teatud protsent inimesi sündinud hormoonidega seotud patoloogiaga - seksuaalse arengu häiretega (NDP) ning nende populatsioonis on võimalik uurida hormonaalse tasakaalutuse seost käitumisega. Alustuseks peaksime lühidalt loetlema emakasisese hormonaalse toime peamised punktid.
Arvatakse, et kõige suurem hormonaalse keskkonna reageerimise periood ilmneb loote küpsemise ajal. Näiteks on teada, et testosterooni maksimaalne toime isaslootele avaldub 8 kuni 24 nädalani ja kordub seejärel sünnist kuni umbes kolme kuuni (Hines xnumx) Kogu küpsemisperioodi vältel pärinevad östrogeenid platsentast ja ema vereringesüsteemist (Albrecht 2010) Loomkatsed näitavad, et erinevate hormoonide suhtes võib olla tundlikkuse perioode palju, ühe hormooni olemasolu võib mõjutada teise hormooni toimet ja nende hormoonide retseptorite tundlikkus võib mõjutada nende tegevust (Berenbaum Xnumx) Loote seksuaalne eristamine iseenesest on uskumatult keeruline süsteem.
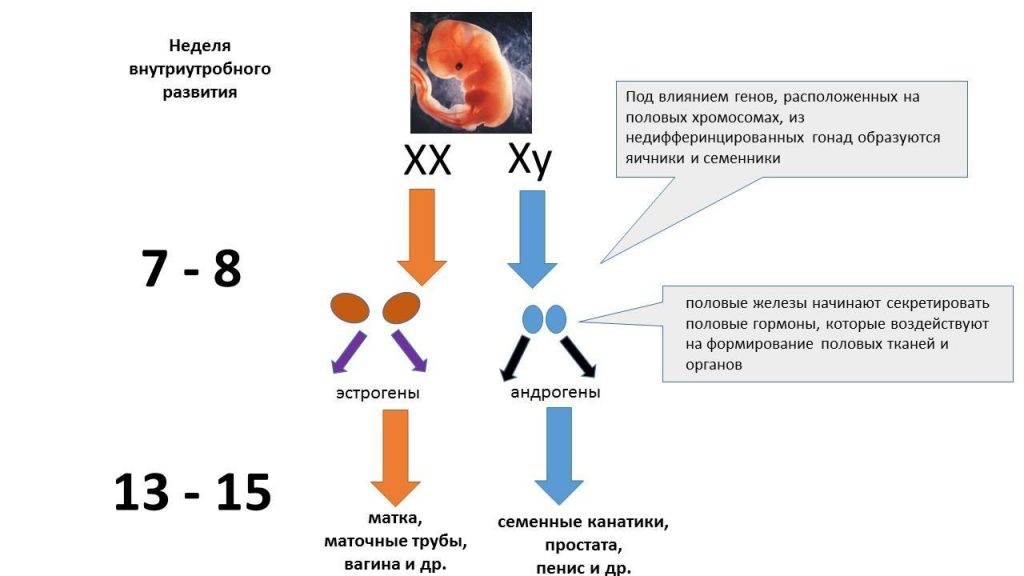
Selles uurimisvaldkonnas pakuvad erilist huvi hormoonid nagu testosteroon, dihüdrotestosteroon (testosterooni metaboliit ja tugevam kui testosteroon), östradiool, progesteroon ja kortisool. Seda peetakse normaalseks, kui hormonaalne mõju loote arengule emakas toimub etapiviisiliselt. Alguses erinevad embrüod ainult nende kromosoomi koostises - XX või XY - ja nende sugu näärmed (sugunäärmed) on samad. Kuid üsna kiiresti, sõltuvalt kromosomaalsest kombinatsioonist, algab munandite (munandite) moodustumine XY kandjatel ja munasarjade tekkimisel XX kandjatel. Niipea kui sugunäärmete diferentseerumine on möödas, hakkavad nad tootma soo-spetsiifilisi hormoone, mis määravad väliste suguelundite arengu ja moodustumise: munandite eritavad androgeenid aitavad kaasa meeste väliste suguelundite arengule ning androgeenide ja naiste östrogeeni puudumine viib naiste väliste suguelundite arengusse. (Wilson 1981).
Seksuaalse eristamise skeem. Koostanud V. Lysov Androgeenide ja östrogeenide tasakaalu rikkumine (geneetiliste mutatsioonide ja muude mõjutuste tõttu), samuti nende olemasolu või puudumine loote teatud olulistel arenguperioodidel, võib põhjustada seksuaalse arengu häireid.
Üks seksuaalse arengu kõige põhjalikumalt uuritud häireid on neerupealise koore kaasasündinud hüperplaasia (VGKN), mis on seotud hormooni kortisooli sünteesis osaleva ensüümi kodeeriva geeni mutatsiooniga (Speiser 2003) See patoloogia põhjustab kortisooli eellaste ülemäärase esinemise (kortisool ja androgeenid jagavad ühist eelkäijat), millest moodustuvad androgeenid. Selle tulemusel sünnivad tüdrukud erineva viriliseerumisastmega8 suguelundid - sõltuvalt geneetilise defekti raskusest ja androgeenide liigsuse astmest. Raskete virilisatsioonijuhtude ja sügavate funktsionaalsete defektide tekkimisega on mõnikord vaja kirurgilist sekkumist. Liigse androgeenide mõju neutraliseerimiseks on ette nähtud hormoonravi. Märgiti, et HCV-ga naistel on suurem risk homoseksuaalse külgetõmbe tekkimiseks (Speiser 2009) ja need, kes kannatasid HCV all raskemal kujul, muutuvad tõenäolisemalt heteroseksuaalseks kui naised, kellel on haigus olnud kergemal kujul (Hines xnumx).
Lisaks on geneetilistel meestel häiritud seksuaalne areng, kes kannatavad androgeenide tundlikkuse puudumise all. Androgeenitundlikkuse sündroomiga meestel toodavad munandid tavaliselt androgeeni testosterooni, kuid testosterooni retseptorid ei toimi. Sündides näevad suguelundid välja nagu naised ja laps on üles kasvanud tüdrukuna. Lapse endogeenne testosteroon muundatakse östrogeeniks, nii et see hakkab arendama naise sekundaarseid seksuaalomadusi (Hughes xnumx) Patoloogia tuvastatakse alles siis, kui puberteet on saavutatud, kui menstruatsioon vastupidiselt ei alga ja muidugi on sellised “naised” viljatud, nagu näiteks viljatus ja “mehed” VGKN-iga.
Mõningaid geneetilisi mehi (s.o XY genotüübiga isikuid) mõjutavaid seksuaalseid funktsioonihäireid on ka teisi, kelle androgeenide puudus tuleneb otseselt ensüümide puudumisest, mis osalevad kas testosterooni dihüdrotestosterooni sünteesis või hormooni eelkäija testosterooni tootmises. Selliste häiretega isikud sünnivad erineva raskusastmega suguelundite deformatsioonidega (Cohen-Kettenis 2005).
Ilmselt seostatakse nendes näidetes homoseksuaalset külgetõmmet ja / või vastassoost lähtuva käitumise valimist funktsionaalsete ja morfoloogiliste patoloogiatega. Kuid selliseid patoloogiaid homoseksuaalidel ei tuvastata. Eeldust, et hormonaalne tasakaalutus mingil moel viib ainult homoseksuaalse eelistuse kujunemiseni (see tähendab, et see mõjutab käitumisjooni) ega mõjuta mingil moel morfoloogilisi ja funktsionaalseid omadusi, empiirilised vaatlused ei toeta.
Homoseksuaalsete eelistustega seotud anatoomilisi ja funktsionaalseid tunnuseid on tehtud mitmesuguseid katseid. Mõelge LGBT + aktivistide viidatud uuringutele.
Simon Levey uurimus
Neurobioloogiliste erinevuste uurimiseks sõltuvalt seksuaalsest kalduvusest on tehtud mitmeid uuringuid. Esimene oli neuroteadlase Simon LeVay avaldamine 1991-is (LeVay 1991). LeVay uuris surnud inimeste lahkamise tulemusi. Ta jagas katsealused kolme rühma - 6 "heteroseksuaalset" naist, 19 "homoseksuaalset" meest, kes surid AIDSi, ja 16 "heteroseksuaalset" meest (need parameetrid on toodud jutumärkides, kuna lahkunu seksuaalsed eelistused olid suures osas spekulatiivsed).
Igas rühmas mõõtis LeVey aju spetsiaalse piirkonna suurust, mida nimetatakse eesmise hüpotalamuse interstitsiaalseks tuumaks.9. Hüpotalamuses eristatakse mitut sellist tuuma suurusega 0.05 kuni 0.3 mm³ (Byne xnumx), mis on nummerdatud: 1, 2, 3, 4. Tavaliselt sõltub INAH-3 suurus meessuguhormooni testosterooni tasemest organismis: mida rohkem testosterooni, seda suurem on INAH-3. LeVey kinnitas, et INAH-3 suurus oli homoseksuaalidel palju väiksem kui meestel, kes olid huvitatud vastassoost, umbes sama palju kui naistel. Kuna inimkeha struktuuri määravad geenid, soovitas LeVey, et kui INAH-3 suurus korreleerub seksuaalse iha suunaga, siis "... libiido on tingitud aju struktuurist ..." ja seetõttu korreleeruvad geenid sugutungiga.
Tuleb märkida, et LeVey pühendus sellele tööle täielikult ja lootis väga saada just selline tulemus. Pärast tema homoseksuaalse partneri Richard Sherry surma aidsisse sattumist oli LeVey mõnda aega depressioonis (Newsweek xnumx, lk 49). Ta ütles ajakirjanikele pärast selle avaldamist, et ta tegi sõna: "Ma tundsin, et kui ma midagi ei leia, loobun täielikult teadusest" (Newsweek xnumx, lk 49).
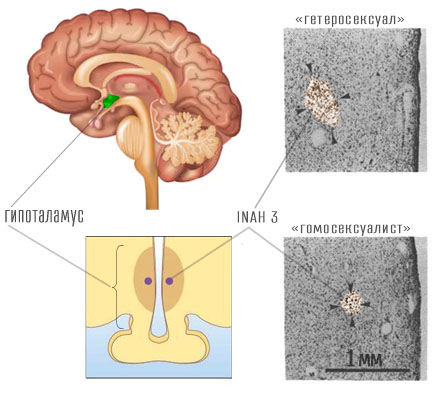
LeVey uurimuses oli palju metodoloogilisi vigu, mida ta ise pidi korduvalt välja tooma, kuid meedia ignoreeris neid kangekaelselt. Mida LeVey tegelikult avastas või ei leidnud? Mida ta ei leidnud ühemõtteliselt, on seos INAH-3 suuruse ja seksuaalsete kalduvuste vahel. Juba 1994-ile allus New Yorgi teadlane William Byne homoseksuaalsuse geneetilise põhjuse kohta avalduse tõsisele kriitilisele analüüsile (Byne xnumx): esiteks on see uurimisobjektide valimise probleem. LeVey ei teadnud täpselt, millised olid seksuaalsed kalduvused inimestel, keda ta oma elu jooksul uuris. On hästi teada, et terminaalse AIDSiga patsientidel täheldatakse madalat testosterooni taset nii haiguse mõju kui ka ravi kõrvaltoimete tõttu (Gomes 2016) LeVay andmete põhjal on täiesti võimatu kindlaks teha, kui suur INAH-3 oli sündides, ja välistada asjaolu, et see võib elu jooksul väheneda. Kõik LeVay poolt homoseksuaalidena tuvastatud isikud surid AIDS-i tüsistustesse. LeVey teeb samas artiklis reservatsiooni:
"... tulemused ei võimalda meil järeldada, kas INAH 3 suurus on inimese seksuaalse sättumuse põhjus või tagajärg või kas INAH 3 suurus ja seksuaalne sättumus muutuvad vastastikku mõne kolmanda tuvastamata muutuja mõjul ..." (LeVay 1991, lk 1036).
Teiseks pole põhjust kindlalt väita, et LeVey avastas üldse midagi. Teadlased Ruth Hubbard ja Elijah Wald oma raamatus Geenide müüdi hävitamine: kuidas teadlased, arstid, tööandjad, kindlustusettevõtted, koolitajad ja inimõiguste kaitsjad manipuleerivad geneetilist teavet, kahtlesid mitte ainult LeVey tulemuste tõlgendamises, vaid ka selles, et kõik olulised erinevused (Hubbard xnumx, lk 95). Ehkki LeVey juhtis tähelepanu sellele, et nende isikute rühmas, keda ta pidas homoseksuaalseteks, oli INAH-3i keskmine suurus väiksem kui INAH-3i keskmisel osal nende rühmas, keda ta pidas heteroseksuaalseteks meesteks, järeldub tema tulemustest, et väärtuste maksimaalne ja minimaalne hajuvus on ideaalselt sama mõlemas rühmas. On olemas statistiline mõiste - normaaljaotuse seadus. Lihtsustatult on selle seadusega öeldud, et atribuudi omanike suurimal arvul on selle atribuudi parameetrid keskmises vahemikus ja ainult vähestel omanikel on äärmise väärtuse parameetrid. See tähendab, et 100 inimestest kasvab 80 160 - 180, 10 on väiksem kui 160, 10 on suurem kui 180 cm.
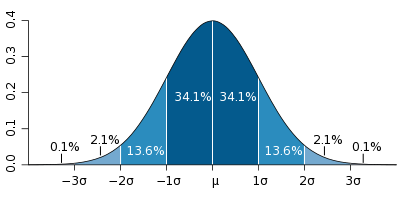
Statistiliste arvutuste reeglite kohaselt on kahe subjektide rühma vahel statistiliselt olulise erinevuse tuvastamiseks võimatu võrrelda parameetrit, millel pole normaalset jaotust. Näiteks kui ühes rühmas alla 160 cm ei ole 10%, vaid 40% või 50%. LeVay uuringus oli INAH-3 väikseim suurus mõne heteroseksuaalse mehe ja enamiku homoseksuaalide jaoks ning maksimaalne suurus mõne homoseksuaali ja enamiku heteroseksuaalsete meeste puhul. Sellest järeldub, et INAH-3i suuruse ja seksuaalse käitumise seose kohta on iga inimese kohta täiesti võimatu midagi öelda. Isegi kui aju struktuuris oleks veenvalt tõendatud erinevuste olemasolu, oleks nende tähtsus sama, mis avastusel, et sportlaste lihased on suuremad kui tavalistel inimestel. Milliseid järeldusi saame selle fakti põhjal teha? Kas inimesel arenevad spordiga mängides suuremad lihased või tekitab kaasasündinud eelsoodumus suuremate lihaste järele inimese sportlaseks?
Ja kolmandaks, LeVey ei öelnud midagi naiste seksuaalse käitumise ja INAH-3 suhete kohta.

1994-i intervjuus ütles LeVey:
"... On oluline rõhutada, et ma ei ole tõestanud, et homoseksuaalsus on kaasasündinud ega ole leidnud selle geneetilist põhjust. Ma ei ole demonstreerinud, et geid “sünniksid nii” - see on kõige tavalisem viga, mida inimesed minu tööd tõlgendades teevad. Samuti ei leidnud ma ajust “geikeskust” ... Me ei tea, kas sündides leitud erinevused olid olemas või ilmnesid hiljem. Minu töös ei käsitleta küsimust, kas seksuaalne orientatsioon loodi enne sündi ...Nimmons xnumx).
LeVey reservatsioon on väga oluline, kuna iga neuroteaduse spetsialist teab sellist nähtust nagu neuroplastilisus - närvikoe võime muuta inimese elus oma funktsiooni ja struktuuri mitmesuguste käitumuslike tegurite mõjul.
2000-is avaldas grupp Briti teadlasi Londoni taksojuhtide aju-uuringu tulemused (Maguire 2000) Selgus, et taksojuhtide jaoks oli ruumilise koordineerimise eest vastutav ajupiirkond palju suurem kui kontrollrühma isikutel, kes ei töötanud taksojuhina, lisaks sõltus selle jaotise suurus otseselt taksoga töötatud aastate arvust (Maguire 2000) Kui teadlased püüdleksid poliitiliste eesmärkide poole, oleks nad võinud öelda midagi sellist: „Nendele taksojuhtidele tuleb anda parempoolne rool ja igal pool, kus nad töötavad, tasub vasakpoolne rool vahetada parempoolse roolimise vastu - sest nad on ju nii sündinud!“

Praeguseks on kogutud veenvat tõendusmaterjali nii ajukudede üldiselt kui ka hüpotalamuse plastilisuse kasuks (Bains xnumx; Müük 2014; Mainardi 2013; Hatton xnumx; Teodoos 1993) Aju morfoloogia muutused käitumuslike tegurite mõjul (Kolb 1998) Näiteks ajustruktuurid muutuvad pärast rasedus (Hoekzema jt. 2016)kosmoses viibimine (van Ombergen jt. Xnumx) ja pärast regulaarset füüsilist tegevust (Nokia jt. Xnumx).
Seetõttu, kinnitades LeVey enda poolt 1994-aastal öeldud sõnu, on tema 1991-aasta uuringu panus homoseksuaalsuse kaasasündinud olemuse hüpoteesisse null.
LeVay töö täpsem kriitika, aga ka muud neuroanatoomilised hüpoteesid on ajakirjas Current Science (Mbugua 2003).
Levay uurimistöö korratavus
Keegi ei suutnud LeVey tulemusi korrata. Aasta 2001 väljaandes viis New Yorgi teadlaste rühm läbi sarnase uuringu - võrreldi hüpotalamuse samu lõike nagu LeVay uuringus, kuid palju täielikumate andmete ja uuritud uuringu piisava jaotusega (Byne xnumx). Nad ei leidnud INAH-3 suuruse sõltuvust homoseksuaalsusest. Autorid jõudsid järeldusele, et "... ainuüksi INAH 3 mahu põhjal ei saa seksuaalset sättumust usaldusväärselt ennustada ..." (Byne xnumx, lk 91).
Hiljem üritati tuvastada seksuaalsete kalduvuste sõltuvust teistest aju osadest. Ajakirjas 2002 avaldasid psühholoog Lasko ja tema kolleegid uuringu aju teisest osast - eesmisest kommissioonist (Lasco 2002) Näidati, et selles piirkonnas ei ole olulisi erinevusi sõltuvalt soost või seksuaalse iha iseloomust. Muud uuringud, mille eesmärk on tuvastada heteroseksuaalide ja homoseksuaalide aju struktuurilised või funktsionaalsed erinevused nende loomupäraste piirangute tõttu, on peaaegu tähelepandamatud: ajakirjas 2008 võeti mõne nende uuringute tulemused kokku artiklis, mis avaldati ajakirjas Proceedings of the US National Teaduste Akadeemia (Swaab xnumx) Näiteks kasutati ühes uuringus funktsionaalset magnetresonantstomograafiat aju aktiivsuse muutuste mõõtmiseks, kui katsealused näitasid meeste ja naiste fotosid. Leiti, et naise nägu suurendas homoseksuaalsete meeste ja naiste aktiivsust talamuses ja orbitofrontaalses ajukoores, samas kui homoseksuaalsete meeste ja heteroseksuaalsete naiste puhul reageerisid need piirkonnad mehe näole paremini (Kranz 2006) Fakt, et heteroseksuaalsete naiste ja homoseksuaalsete meeste ajud reageerivad eriti meeste nägudele, samal ajal kui heteroseksuaalsete meeste ja homoseksuaalsete naiste ajud reageerivad eriti naiste nägudele, pole homoseksuaalsete kalduvuste etioloogiat arvestades vaevalt suur avastus. Samamoodi tsiteeritakse teises uuringus erinevaid reaktsioone feromoonidele mittehomoseksuaalsetel ja homoseksuaalsetel meestel (Savic 2005).
Sõrme pikkus
Teise sõrme (indeksi) pikkuse ja käte neljanda sõrme (rõnga) pikkuse suhe, mida tavaliselt nimetatakse suhteks “2D: 4D”, on enamiku meeste ja naiste puhul erinev. Mõned tõendid viitavad sellele, et see suhe võib sõltuda emakasisese testosterooni tasemest, mille tulemuseks on kõrgema testosterooni kontsentratsiooniga meestel, nimetissõrm on lühem kui sõrmusesõrm (st 2D: 4D madal suhe) ja vastupidi (Hönekopp 2007) Mõne teadlase sõnul on 2D: 4D indeks seotud homoseksuaalsete kalduvustega. Katsed kuidagi 2D: 4D suhet seostada ja seksuaalsed kalduvused on vastuolulised ja vaieldavad.
Ühe hüpoteesi kohaselt võib homoseksuaalidel olla suurem 2D: 4D suhe (lähemal naiste suhtele kui heteroseksuaalsete meeste suhtele), samas kui teine hüpotees viitab vastupidiselt sellele, et sünnieelse testosterooni abil hüpermaskuliineerimine võib põhjustada madalama suhte homoseksuaalid kui heteroseksuaalsed mehed. Esitati ka hüpotees naiste homoseksuaalsete kalduvuste kohta hüpermaskuliniseerumise tagajärjel (madalam suhe, kõrgem testosterooni tase).

Mitmed võrdlevad uuringud selle tunnuse kohta homoseksuaalsete ja mittehomoseksuaalsete naiste ja meeste osas on andnud erinevaid tulemusi. Ajakirjas Nature ajakirjas 2000 avaldatud uuring näitas, et 720-i täiskasvanud kalifornlaste valimis oli parema käe suhe 2D: 4D samasooliste eelistustega naistel märkimisväärselt mehelikum (s.o madalam) kui mittehomoseksuaalsetel naistel ja märkimisväärselt ei erinenud mittehomoseksuaalsete meeste suhtarvust (Williams 2000) See uuring ei tuvastanud ka olulist erinevust geide ja homoseksuaalide keskmiste 2D: 4D suhete vahel. Samal aastal näitas teine uuring, kus kasutati suhteliselt väikest Suurbritannia homoseksuaalsete ja mittehomoseksuaalsete meeste valimit, 2D madalamat väärtust: 4D (s.o mehelikum) homoseksuaalide hulgas (Robinson 2000) 2003 aastal leidis londonlaste valimi uuring, et homoseksuaalide 2D: 4D määr oli madalam kui mittehomoseksuaalsete meestega (Rahman xnumx), samal ajal kui kaks muud Californias ja Texases asuva proovi uuringut näitasid 2D kõrgemat väärtust: 4D homoseksuaalide jaoks (Lippa xnumx; McFadden 2002) 2003-is viidi läbi võrdlev uuring seitsme paari monosügootsete kaksikute naiste kohta, kõigis paarides oli ühel kaksikutest naistel homoseksuaalne eelistus ja viis paari monosügootseid kaksikute naisi, kus mõlemal õel olid samast soost eelistused (Saal 2003) Erinevat tüüpi seksuaalse külgetõmbega kaksikute paaris oli inimestel, kes identifitseerivad end homoseksuaalseks, suhe 2D: 4D oluliselt madalam kui nende kaksikute oma, samal ajal kui samalaadsed kaksikud ei leidnud erinevust. Autorid järeldasid, et see tulemus osutab, et "2D: 4D madal suhe on sünnieelse keskkonna erinevuste tulemus". Ja lõpuks leiti 2005 aastal Austria 2 homoseksuaalsete meeste ja 4 mitte-gei meeste valimis 95D: 79D suhte uuringu tulemusel, et indikaatorite 2D: 4D mitte-geide puhul ei erinenud oluliselt homoseksuaalsete meeste näitajatest (Voracek 2005) Pärast selle tunnuse mitmete uuringute ülevaatamist järeldavad autorid, et "on vaja rohkem andmeid, et saaksime enesekindlalt järeldada, kas suhte 2D: 4D ja meeste seksuaalse iha olemuse vahel on seos, arvestades etnilisi erinevusi".
Silm pilgutab
In 2003 teatas grupp inglise teadlasi, et nad on leidnud "uusi veenvaid tõendeid selle kohta, et seksuaalne iha on tingitud inimese aju omadustest" (Rahman xnumx) Katsi Rahman ja kaasautorid leidsid, et vastusena valjule mürale leidsid nad erinevat reaktsioonikiirust - silmade pilgutamist. Autorid leidsid, et naistel on vähem nn "Impulsi-eelne pärssimine" (PPI) - keha motoorse vastuse langus stiimulitele nõrga eelstimendi juuresolekul10... See tähendab, et naised pilgutasid kiiremini kui mehed ja samasooliste eelistustega naised aeglasemalt kui mittehomoseksuaalid. Tuleb märkida, et esiteks viisid autorid läbi uuringu väikeses uuritavate rühmas ja teiseks ei leidnud nad erinevusi homoseksuaalsete meeste ja mittehomoseksuaalsete meeste vahel. Sellest hoolimata otsustasid autorid, et nende tulemused tõestavad, et homoseksuaalsus on kaasasündinud nähtus. Sellegipoolest tegid teadlased siiski mõned reservatsioonid: nad märkisid, et küsimus, kas leitud erinevused tulenevad seksuaalse külgetõmbe eripärast või tulenevad konkreetsest seksuaalkäitumisest, on endiselt lahendamata. Nad tõid välja: "... heteroseksuaalide ja homoseksuaalide neuroanatoomilised ja neurofüsioloogilised erinevused võivad olla tingitud kas bioloogilistest teguritest või õppimise mõjust ...". Dr Halstead Harrison Washingtoni ülikoolist analüüsis seda uuringut ja märkis sellise olulise puuduse nagu testitud rühmade väike suurus (14 homoseksuaalset naist ja 15 heteroseksuaalset naist, 15 homoseksuaalset meest ja 15 heteroseksuaalset meest). Harrison järeldas: "Rahman jt. Ei esitanud veenvaid tõendeid järelduse toetamiseks, et homoseksuaalsete naiste PPI parameetrid sarnanevad meeste omadega."Harrison xnumx) Harrison seadis kahtluse alla ka meetodite statistilise adekvaatsuse.
Eespool käsitletud kaksikute uuringud võivad selgitada emahormoonide mõju määra, kuna emakasisese arengu ajal kogevad identsed ja identsed kaksikud oma mõju samal viisil. Kaksikute uuringute nõrgad vastavusnäitajad näitavad, et sünnieelsed hormoonid kui geneetilised tegurid ei mängi määravat rolli seksuaalse iha tekkimisel. Ka muud katsed leida seksuaalset iha oluliselt mõjutavaid hormonaalseid tegureid on olnud ebaselged ja nende tulemuste olulisust pole veel mõistetud.
Ema stressi tagajärjed
1983-is viisid Gunther Dörner jt läbi uuringu, et teha kindlaks seos ema raseduse ajal tekkiva stressi ja nende laste järgneva seksuaalse identiteedi vahel. Nad küsitlesid kakssada inimest sündmustest, mis võivad nende emadel raseduse ajal stressi põhjustada - st vastajate endi emakasisese arengu (Dörner 1983) Paljud sündmused olid seotud II maailmasõja tagajärgedega. Meestest, kes teatasid, et nende emad kogesid raseduse ajal mõõdukat kuni tugevat stressi, olid 65% homoseksuaalsed, 25% olid biseksuaalsed ja 10% olid heteroseksuaalsed. Hilisemates uuringutes täheldati siiski kas palju väiksemaid korrelatsioone või oluliste korrelatsioonide puudumist (Ellis 1988) Pärast 2002is läbi viidud uuringut, mis käsitles sugutungi ja sünnieelse stressi vahelist seost teisel ja kolmandal trimestril, leidsid Hines ja tema kolleegid, et raseduse ajal olnud ema stress oli "vaid pisut seotud" nende tütarde tüüpilise meeskäitumisega 42-kuuste vanuselt ". ja puudub igasugune seos ”oma poegade tavaliselt naiseliku käitumisega (Hines xnumx).
Kolmas osa: immuunsüsteemi häired?
Suure venna efekt
„Vanema venna mõju” (ESB) või „vendade sünnijärjekorra mõju”11 - selle termini pakkusid välja Kanada-Ameerika teadlased Ray Blanchard ja Anthony Bogert - see on see, et mõnede tähelepanekute kohaselt on homoseksuaalsetel pedofiilidel, homoseksuaalidel ja vägistajatel võrreldes tavaliste heteroseksuaalsete meestega rohkem vanemaid vendi, kuid mitte vanemaid õdesid (Blanchard 1996; Bogaert 1997; Blanchard 1998; Lalumiere 1998; Blanchard 2000; 2002i hinnang; MacCulloch 2004; Blanchard 2018).

Praegu on avatud arutelu selle üle, kas (1), kas ESB on tõesti olemas, ja (2), kui see on olemas, kas sellel on bioloogiline või sotsiaalne põhjus (Zietsch 2018; Gavrilets 2017; Valgepea 2018).
Vaatamata vastuolulistele tulemustele ESB ja selle põhjuste vallas, võtsid mõned teadlased ja avaliku elu tegelased, püüdes leida homoseksuaalsusele bioloogilisi põhjendusi, ESB bioloogilise seletuse nii selgelt omaks, et välistasid täielikult kõik muud võimalikud seletused (kasvatuse mõju jne). .).
🇧🇷2023 lisa:
Vilsmeier JK, Kossmeier M, Voracek M, Tran US. 2023. Vennaliku sünnijärje efekt kui statistiline artefakt: koonduvad tõendid tõenäosusarvutusest, simuleeritud andmed ja mitmekülgne metaanalüüs. Peer J 11:e15623 https://doi.org/10.7717/peerj.15623
Viini ülikooli psühholoogiaosakonna teadlased viisid läbi suure venna efekti andmete matemaatilise töötlemise. Nad jõudsid järeldusele, et kui seda õigesti analüüsida, on vanemate vendade arvu ja homoseksuaalse orientatsiooni vaheline konkreetne seos väike, heterogeenne ega ilmselt meestele omane. Lisaks olemasolevad teaduslikud tõendid liialdatud väikeste uuringute mõju tõttu.
ESB hüpoteesi puudused
ESB ei ole tingimusteta aksioom, selle olemasolu on paljudel põhjustel pideva teadusliku arutelu objekt.
Esiteks ei tuvastata seda mõju kõigis uuringutes. Brendan P. Zietsch märkis, et ESB hüpoteesi toetajad lisavad oma analüüsidesse ainult avaldatud uuringute tulemusi, mis vastavad nende ideedele, ning jätavad tähelepanuta uuringud, infolehed, väitekirjad ja konverentsidel esinevad ettekanded, kus ESB-d ei tuvastata (Zietsch 2018) See probleem on eriti oluline, arvestades, et kuues seitsmest õigesti sarnasest tõenäosusvalimist ei kinnitatud ESB-d (Bearman 2002; Bogaert 2005, 2010; Francis xnumx; Frisch xnumx; Zietsch 2012) Eelpool mainitud Simon LeVay liikumise LGBT + aktivist annab oma töös ülevaate ka uuringutest, milles ESB-d ei tuvastatud (LeVay 2016).
Teiseks, need uuringud, milles ESB tuvastati, põhinevad kahtlasel proovivõtumeetodil. ESB hüpoteesi toetajad rakendavad populatsiooni analüüsimisel selliseid kriteeriume, mis välistavad kõigi saadaolevate tõenäosuslike valimite (s.o need valimid, mis valitakse juhuslikult uuritud sõltumatu muutuja - seksuaalse külgetõmbe - suhtes). See tähendab, et metaanalüüs hõlmab ainult neid proove, milles homoseksuaalide osakaal ei sarnane homoseksuaalide osakaaluga elanikkonnast (näiteks aasta 2018i Blanchardi analüüsi proovid sisaldavad keskmiselt 51% homoseksuaalidest, samas kui nende elanikkonna üldpopulatsioonis) erinevate allikate kohaselt on maksimaalne väärtus 2 - 3%). Selliste mitte-juhuslike proovide korral suureneb homoseksuaalsete ja heteroseksuaalsete rühmade valimise oht, mis erinevad mitte ainult ennustaja muutujate osas. Blanchard 1i tabel 2018 näitab, et suurem osa meta-analüüsi kaasatud proovidest on võetud äärmiselt esindamatutest populatsioonidest: seksuaalkurjategijad, transsoolised inimesed, pedofiilid, psühhopaadid jne. On tähelepanuväärne, et ühtegi neist valimi valimise probleemidest artiklis ei käsitletud. Vastupidi, Blanchardi kaasamiskriteeriume rakendati nii, et välistati suured uuringud tõenäosusvalimitega (mille puhul ESB-d ei kinnitatud). Metaanalüüsi üksikute uuringute vahelise mõju suur heterogeensus näitab, et see, kuidas rühmad uuringuks valitakse, mõjutab suuresti ESB-d. See suurendab tõenäosust, et valimi omadused loovad ESB, eriti kui arvestada, et suure tõenäosusega valimid ei näita üldse ESB-d.
Kolmandaks, teine metoodiline probleem on see, et ESBde leidmise analüütilised meetodid näivad olevat erapoolikud ja suunatud soovitud efekti tuvastamisele. Näiteks kasutasid mõned teadlased mõju mõõtmiseks ühesuunalist statistilist testi (nt Bogaert 2005; Poasa 2004; Purcell 2000) või tõlgendanud teiste teadlaste tulemusi, kes ei tuvastanud ESB-d märkimisväärselt, öeldes, et oleks tulnud kasutada ühesuunalisi teste (Blanchard 2015) - kuigi on teada, et ühesuunalisi teste saab kasutada ainult väga harvadel juhtudel, mis ei sobi meta-analüüsi tingimustele (Lombardi xnumx) Teadlane Bartlett kirjutab järgmist:
"... Arvestades homoseksuaalsete meeste suhtelist nappust elanikkonnas, on uuringu jaoks keeruline leida tasakaalustatud homoseksuaalide ja heteroseksuaalsete meeste rühmi. ESB mõõtmisel on probleemiks erineva perekonna suurusega populatsioonidest pärit homoseksuaalide ja heteroseksuaalide valimine. Tõenäosus, et uuring leiab võltsmõju kõikvõimalike õdede-vendade, mitte ainult vanemate õdede-vendade jaoks, suureneb, kui valimisse kuuluvad suuremate perede homoseksuaalid, samas kui efekt kaob, kui valimisse kuuluvad heteroseksuaalsed mehed suurematest perekondadest. ... "(Bartlett xnumx).
Neljandaks põhineb ESB üksnes korrelatsioonianalüüsi tulemustel. Tegelike korrelatsioonide tuvastamine on identne selle korrelatsiooni loonud põhjuse tuvastamisega. Mis tahes korrelatsioonid vajavad ka mehhaanilist selgitust selle kohta, mida ei täidetud (Gavrilets 2017).

Viiendaks, ESB pole universaalne. ESB ei suuda selgitada homoseksuaalsust meestel, kellel pole vanemaid vendi, samuti ei suuda ta selgitada homoseksuaalse külgetõmbejõudu noorematel vendadel, kellel on homoseksuaalsuse vanem vend, ei suuda selgitada kaksikvendade seksuaalsete eelistuste lahknevust12. ESB ei esine biseksuaalsetel meestel. Biseksuaalset külgetõmmet võib mõista kui seksuaalset külgetõmmet nii vastupidise kui ka oma soo suhtes, seetõttu peaks ESB paradigma raames biseksuaalsetel meestel olema vähem ESB kui homoseksuaalsetel meestel, kuid rohkem kui heteroseksuaalsetel meestel. Uuringus siiski Bogaert (2006) ESB oli sama nii biseksuaalsete kui ka homoseksuaalsete isikute puhul. McConaghy ja tema kolleegid (2006) viis ESB uuringu läbi valdavalt heteroseksuaalsete isikutega (indiviidid, kellel on kerge samasoolisus), võrreldes erandlike heteroseksuaalide kontrollrühmaga. ESB-d on täheldatud nii meeste kui ka naiste puhul. Lisaks täheldati vanema õe mõju ka meestel, ehkki vähem tugev. Autorite sõnul näitavad nende tulemused, et ESB bioloogilised põhjused on vähem tõenäolised kui sotsiaalsed. Hinnanguliselt selgitab ESB hüpotees ainult 17% homoseksuaalse külgetõmbejuhtumite koguarvust ja ainult meestel (Cantor xnumx) ESB ei selgita naiste homoseksuaalseid eelistusi. ESB hüpoteesi toetajad on mitu korda proovinud seda efekti leida homoseksuaalsete eelistustega naistel, kuid tulemusteta (Blanchard 2004).
Kuuendaks, ESB ei tööta tegelikes kultuurilis-etnilistes ennustusmudelites. Eeldades, et ESB eksisteerib vastavalt selle paradigmale, võib ennustada (mudel vastavalt Bogaert 2004), et homoseksuaalsete eelistustega meeste seas on suur esinemissagedus: a) usuperedes, kus suure hulga laste tõenäosus on suurem; c) ida- ja moslemikultuur, mida tavaliselt eristavad suured pered; ja madalam levimus - kõrge elatustasemega lääne ühiskondades, kus sündimus on oluliselt madalam kui ida ühiskondades (Caldwell 1997) Sarnane suundumus, pehmelt öeldes, ei vasta tegelikkusele.
ESB hüpoteesid
Mõnedes uuringutes leitud ESB-d selgitavad mitmed eeldused (James xnumx), nende hulgast saab eristada kahte peamist: (1) sünnieelne bioloogiline kokkupuude (ema immuniseerimise hüpotees) ja (2) sotsiaalselt psühholoogiline sünnitusjärgne (kokkupuude keskkonnatingimustega). Allpool analüüsime mõlemat eeldust.
Ema immuniseerimise hüpotees
Blanchard ja Bogert esitasid ESB bioloogilise alusena hüpoteesi ema immuunkonflikti kohta, mille kohaselt naise immuunsüsteem on väidetavalt võimeline tootma isasloote teatud "mehaaniliste antigeenide" vastu antikehi ning väidetavalt sarnaseid antikehi koguneb isasloote iga järgneva raseduse ajal, iga järgmise poisi emakasisese immuunkahjustuse riski suurendamine (Blanchard 1996) Ema immuunkonflikti hüpotees püüab poisi homoseksuaalsete eelistuste arengut selgitada analoogia põhjal Rh-konfliktiga raseduse ajal (Bogaert 2011).
Reesuskonfliktiline rasedus on patoloogiline seisund, mis on põhjustatud vererakkudel spetsiifilist valku kodeeriva geeni olemasolust loos ja sellise geeni puudumisest emal (see tähendab, et selles näites on ema Rh-negatiivne ja loode Rh-positiivne). Rh-positiivse lootega Rh-negatiivse ema esimese raseduse ajal tungivad loote rakud ema vereringesse ja põhjustavad immuunreaktsiooni - vererakkude antikehade moodustumist. Selle Rh-positiivse lootega ema hilisemates rasedustes tungivad ema vereringes olevad antikehad loote verre ja hävitavad selle punased verelibled, põhjustades sünnil hemolüüsi ja ikterust. Sellepärast kontrollivad sünnitusabi-günekoloogid rase ema ja lapse isa Rh staatust.
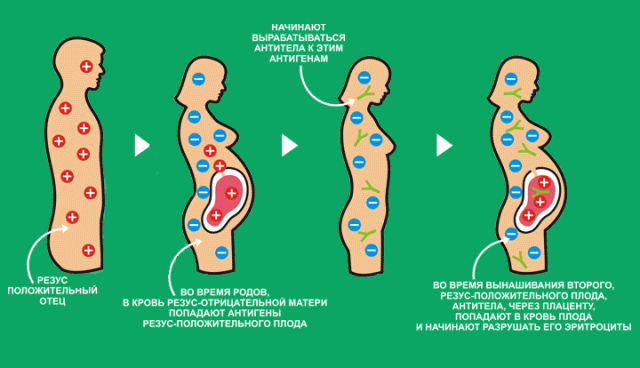
Blanchardi ja Bogerti hüpotees põhineb samadel põhimõtetel nagu Rh-konfliktiga rasedus. Sel juhul on antikehade moodustumist põhjustavaks teguriks (ülaltoodud näites Rh-positiivsus) mängukromosoomi olemasolu, see tähendab loote meessugu. Y-kromosoom kodeerib valkude ja hormoonide moodustumist, mis esinevad isaslootel (kuid mitte naissoost!) Juba embrüogeneesi kõige varasemas etapis. Arutatud hüpoteesi kohaselt sisenevad lootekoe osakesed, mis kannavad “meessogeeni antigeeni”, ema vereringesse ja põhjustavad antikehade teket, mis väidetavalt ületavad hematoentsefaalbarjääri meessoost loote hilisemate raseduste ajal, tungivad loote ajusse ja ründavad spetsiifilisi “meessoost antigeeni” sisaldavaid närvirakke. ", Väidetavalt embrüonaalse aju arengu takistamine" meessoost tüübi poolt ", mille tagajärjel poiss sünnib koos" naiseajuga "ja arvatavasti muutub homoseksuaalseks või transsooliseks. Ema immuunreaktiivsus suureneb isasloote iga uue rasedusega, seetõttu suureneb väidetavalt kõrvalekallete tõenäosus iga vanema venna puhul.
Blanchardi ja Bogerti hüpoteesi kohaselt on emakasisese immuunkahjustuse kinnitus kehakaalu langus sündides homoseksuaalsetel meestel, kellel on vanemad vennad.
Ema immuniseerimise hüpoteesi puudused
William H. James (2004) uuris kriitiliselt ema immuunkonflikti hüpoteesi põhiprintsiipe.
Esiteks on kahtlane eeldus, et raseduse ajal immuniseeritakse ema ainult meessoost loote spetsiifiliste antigeenidega, kuid mitte emasloomadega - pehmelt öeldes. Emad võivad tekitada nii meeste kui ka naiste lootele immuunreaktsioone, see tähendab, et mitte “meessoost antigeenid”, kuid spetsiifilistel isadel on neil juhtudel immuunreaktsioonivõime ja selliseid patoloogiaid on hästi uuritud (Tantsijad xnumx) Kõige tavalisemad on kolm sellist reaktsiooni: a) eelnimetatud RCH, milles mõjutavad loote punaseid vereliblesid ja mille pinnal on positiivne Rh-faktor, sagedus 10 - 20%; b) trombotsüüte mõjutavate vastsündinute alloimmuunne trombotsütopeenia, esinemissagedus 4% või 12%, kui võtta arvesse ka asümptomaatilist vormi (Pööre 2005); vastsündinute neutropeenia, mõjutab neutrofiile, sagedus 4% (Han 2006) Kõigil neil juhtudel on antigeenid isalised, mitte tavalised isased. Nad arenevad samast isast järgmiste sugupoolte lastena. Need mõjutavad verekomponente (ja mitte teatud elundeid ja kudesid) lootevere (nabanööri, platsenta jt) kokkupuutel ema immuunsussüsteemiga (väliste suguelundite, emaka sisepinna jms trauma tõttu) sünnituse ajal.
Ema alloimmuunsed antikehad tungivad väidetavalt ema piima, nagu ka kõik muud antikehad (Gasparoni xnumx), näiteks Rh-faktori alloimmuunsed ema antikehad, mis tungivad ema piima, võivad põhjustada vastsündinu hemolüütilist haigust (Õlu 1975) Sarnaselt võib eeldada, et hilisemad vennad taluvad „meessoost antigeenide“ hüpoteetilisi antikehi sisaldavat piima halvasti, mis põhjustab probleeme rinnaga toitmise ja selle varajase lõpetamisega, samuti allergilise koliidiga. Meditsiinikirjanduse ülevaade annab aga täiesti vastupidise pildi: sünnijärjestus ei ole seotud imetamise kestusega ega korreleeru sellega üldiselt positiivselt (Martin 2002) Allergilise koliidi esinemissagedus vastsündinutel on vahemikus 0,01% kuni 7,5% (Hildebrand xnumx; Pumberger xnumx; Xanthakos 2005), samal ajal mõjutavad mõlemast soost vastsündinud. Sellesse statistikasse kuuluvad ka reaktsioonid lehmapiimale.
Kordame, et evolutsiooni seisukohast on isase loote emakasisene immunogeensus ema jaoks absurdne. Inimese fülogenees imetajana kestab mitu miljonit aastat. Miks pole inimkehas nii pikka aega välja töötatud tõhusaid viise immuunvastuste kujunemise seisukohalt nii kulukate vältimiseks? Naiste keha hüpoteetilised immuunvastused terve naise keha jaoks evolutsiooniliselt nii rutiinse ja vältimatu protsessi ajal, kui rasedus isase lootega, mis moodustab 50% kõigist rasedustest, põhjustaks olulist soolist tasakaalustamatust ja evolutsioonilisi probleeme. Fülogenees viib alati liigi jaoks kõige optimaalsemate tunnuste valimise ja säilitamiseni. Näiteks on olulisi tõendeid selle kohta, et meessoost partneri valik on seotud suurema histo-ühilduvuskompleksiga (GCS) (Chaix 2008; Millinski 2006; Wedekind xnumx), see tähendab, et fülogeneetilisel tasandil on liigiprotsessid maksimaalselt suunatud mitmekesisuse suurendamisele GCS alusel ja järglaste elujõulisuse suurendamisele (Williams 2012; Guleria 2007).
Bogert toob oma teooria kaitsmiseks näitena sellise patoloogilise immuunvastuse nagu Rh-konfliktiga rasedus (RCH) (Bogaert 2011), mis põhjustab vastsündinu hemolüütilist haigust - väidetavalt on see nähtus (risk on umbes 15% elanikkonnast (Izetbegovic 2013)) ei kadunud evolutsiooni käigus. Siiski tuleb meeles pidada, et FC esinemissagedus inimkonna kui liigi minevikus oli oluliselt madalam. Praeguses etapis täheldatakse sellist evolutsioonifaktorit nagu inimkonna segadus, mistõttu ei tundu paradoksaalne, et Rhesuse konflikti blokeerimise loomulikud mehhanismid pole veel välja kujunenud. Transplantoloogia arenguga on inimkond kohanud sellist faktorit, mis varem puudus kui immuunsuse hülgamisreaktsioonid (peaaegu 100% -l retsipientidest), pole üllatav, et inimestel puudub loomulik mehhanism nende allasurumiseks. Inimese kui liigi RCH ja siirdamise hülgamisreaktsioonide korral pole kompensatsioonimehhanismide väljatöötamiseks palju aega möödunud13. Teisest küljest oleks emade immuunsuse mitteühilduvuse stabiilne säilitamine 50% -ga nende järglastest paradoksaalne.
Üldiselt tundub kaheldav, kas meessoost lootel on teatud struktuure või aineid, millel on antigeensed omadused ainult meestele. Vaba testosteroon, suguhormoone siduva globuliini või rakumembraani androgeeniretseptor, ei ole ema suhtes immuunreaktiivne, kuna need kõik esinevad ka naisorganismis.
Teiseks, eeldus, et spetsiifilised ema antikehad kahjustavad valikuliselt meessoost loote aju (põhjustades selle “feminiseerumist”), kuid samal ajal ei häiri nad ühtegi muud aju funktsiooni ega mõjuta munandit (sisaldavad palju rohkem Y-kromosoomi geenide tooteid) ) - on pehmelt öeldes vaieldav.
Kui tegelikult ilmnes immuunreaktsioon “meessoost antigeenide” vastu, siis mõjutavad hüpoteetilised ema antikehad peamiselt ja peamiselt või vähemalt samaaegselt munandeid, mis sisaldavad palju rohkem “meessoost antigeeni” kui aju. On teada palju meesspetsiifilisi geene (st asuvad Y-kromosoomis) (Ginalksi xnumx) Nende geenide ekspressioon - see tähendab teabe lugemine ning valkude ja struktuuride süntees - toimub mitte ainult ajus, vaid peamiselt munandites, mis peaks olema “meestevastase” spetsiifilise immuunsuse rünnaku peamine eesmärk, mitte aju (Ginalksi xnumx) Homoseksuaalsetes meestes võib täheldada munandite patoloogiate suurenenud levimust: hüpospadiad, krüptoridism, munandivähk jne, kuid munandihäirete seost homoseksuaalsuse või ESB-ga ei leitud (Pierik xnumx; Flannery xnumx) Lisaks on huvitav märkida, et hüpospadiaga meestel on hoolimata sünnituseelse arengu madalast testosterooni tasemest pisut kõrgem psühholoogiline mehelikkus (Sandberg 1995) Samuti võib eeldada, et homoseksuaalse külgetõmbega inimestel ilmneb puberteet hiljem immuunsete munandite kahjustuste tõttu, kuid suured uuringud ei tuvastanud puberteediea erinevusi sõltuvalt seksuaalsetest eelistustest (Savin-Williams 2006).
Lisaks oleks ema hüpoteetiliste antikehade sisenemine vereringe kaudu loote ajus võimatu vere-aju barjääri (BBB) tõttu, mis moodustub juba 4-ndal rasedusnädalal (Zusman 2004) Sellised antikehad suudaksid BBB-st üle saada ainult viimaste tõsiste patoloogiate korral - kaitsefunktsioonide rikkumisega, mis tooks kaasa aju olulise neuroloogilise kahjustuse. Kui loote BBB on aga normaalses seisundis, ei põhjusta isegi ema immuunsussüsteemi rikkumised vastsündinu neuroloogilisi patoloogiaid - BBB hoiab ära antikehade teket. 17 283 lastega emade paari hõlmavas suures uuringus ei leitud seost ema suurenenud immunoreaktiivsuse ja tserebraalparalüüsi, vaimse alaarengu, krampide jne (Flannery xnumx).
Samuti pole vastuvõetav hüpotees, et hüpoteetilised antikehad kahjustavad aju nii, et need põhjustavad selle feminiseerumist. Embrüogeneesi staadiumis on aju anatoomilised soolised erinevused nõrgalt väljendatud ja aju lõplik morfofunktsionaalne moodustumine vastavalt soole toimub puberteedieas, kui hüpoteetiline immuunne toime on võimatu (Lenroot 2007; Paus xnumx) Idee teatud sugupoolele iseloomuliku neuraalse organisatsiooni embrüo olemasolu kohta ajus on väga kaheldav ja seda pole kunagi veenvalt tõestatud (Lauterbach 2001; Nunez 2003) MRT-skaneeringud näitasid vastsündinute ajustruktuurides vaid ebaolulisi statistilisi kui dihhotoomseid erinevusi, sugude vahel olid olulised vasted (Zanin xnumx; Mitter 2015).
Hüpoteesi kohaselt peaksime eeldama, et vanemate vendadega homoseksuaalid, kellel on “feminiseeritud” aju, kuuluvad alati fenotüüpi, millel on tavaliselt naiste huvid ja käitumine, kuna on äärmiselt spekulatiivne uskuda, et aju “demaskuliniseerumine” mõjutab ainult poisi seksuaalseid eelistusi, kuid möödub teistest konkreetsed meessoost omadused. Tuleb märkida, et mõnedes uuringutes korreleerub samasooliste ligitõmbamine täiskasvanutel rohkem “naissoost” ajustruktuuridega, kuid aju areng toimub suuruse ja funktsiooni osas peamiselt pärast sündi ja seetõttu on sellised struktuurid autorite endi sõnul sünnitusjärgse tulemuse tagajärg kogemus, mitte sünnieelsed tegurid. Bogaert jt uurimused (2003; 2005); Kishida jt (2015); Semenyna jt (2017) ei tuvastanud korrelatsioone ESB ja naiselike märkide raskuse vahel meestel.
Kolmandaks, hüpoteetilise emakasisese immuunsuse kahjustuse, vanemate vendade arvu, homoseksuaalse külgetõmbe ja kaalulanguse vaheline seos sündides on vähemalt kahtlane.
Üldise immuunsuse rünnaku tõendusmaterjalina viitavad ESB hüpoteesi pooldajad ja immuunsuse kahjustused andmetele, et vanemate vendadega meestel oli väiksem sünnikaal (Blanchard 2001) Blanchardi uuringutes oli kehakaalu langus sündides poistel, kellel on vanemad vennad, umbes 170 grammi (5% kehakaalust) (Blanchard 2001) Arutatava hüpoteesi kohaselt tuleks sarnast langust täheldada homoseksuaalsete eelistustega poiste puhul, kellel on vanemad vennad, ja seda ei tohiks täheldada tüdrukute puhul. Kuid see pole nii - Norra uuringus, milles uuriti immuunvastuste ja kaalukaotuse hüpoteetilist suhet sündides, uuriti 181 000 sünnijuhtumeid ja kaalukaotust sündides täheldati nii tüdrukutel kui ka poistel (Magnus 1985) Lisaks täheldati hüpoteetilist “suure venna efekti” mõlemast soost ja see oli äärmiselt madal - 0,6%, väljendatuna 20 ± 4,5 grammi erinevusena võrreldes normaalse sünnikaaluga 3 500 grammides (Magnus 1985).
Nende andmete kohaselt näib immuunfaktorite roll kehakaalu vähendamisel üldiselt kaheldav. On tähelepanuväärne, et Magnus ja tema kolleegid uurisid ka isapoolsete antigeenide mõju vastsündinute kehakaalule - sel juhul tehti ettepanek, et kui kaalulangus on põhjustatud isapoolsete antigeenide immuunvastustest, märgitakse seda nii poistel kui tüdrukutel. Magnus ja kolleegid uurisid mõlemast soost laste kehamassi sündides emadel, kes on sõlminud uue abielu ja sünnitanud uusi lapsi - kui kaalukaotus oli tingitud immuunvastustest, oleks pidanud teise mehe laste sünnikaal olema naasta standardsete algnäitajate juurde, kuna teine isa on uute antigeenide kandja ja immuunsete antikehade kogunemiseks on vajalik progresseeruv immuunprotsess (mitu rasedust) (Magnus 1985) Teisest isast pärit laste kehakaal sündides jäi siiski väiksemaks ja autorid järeldasid, et nende valimis ei kinnitata kõigi immuunprotsesside seost kehakaalu langusega sündides (Magnus 1985).
Sünnituse ajal saadud kaalukaotuse põhjus võib olla: a) enneaegne sündimine; b) platsenta puudulikkus; c) emade autoimmuunhaigused, näiteks süsteemne erütematoosluupus (kombineerituna paljude kaasasündinud patoloogiatega sündides); d) munandihäiretega seotud patoloogiate kompleks. Ühtegi ülaltoodut pole homode puhul, kellel on vanemad vennad, täheldatud.
Sünnitusjärgse kaalukaotuse seost immuunreaktsioonidega ei ole selgitatud ja see on endiselt väga spekulatiivne küsimus. Vastavalt James (2006) kehakaalu märkimisväärne langus sündides võib olla tingitud testosterooni mõjust (Manikkam 2004) Lisaks on suurenenud testosterooni tase naise kehas seotud poisi sünnituse tõenäosusega (James xnumx; James 2004b) Blanchard, kes töötas välja oma hüpoteesi seda toetavate tõendite kvaliteedis, viitas uuringule Gualtieri ja Hicks (1985)kes väitis, et sündinud laste seksuaalne osakaal on naissoost poole nihkunud sõltuvalt laste arvust (ehk teisisõnu: mida rohkem lapsi perre sündis, seda väiksem oli poisi sündimise tõenäosus). Selles uuringus oli siiski tõlgendusviga (vt James xnumx, lk 52; James xnumx) Seevastu kaks suurimat uuringut: 4 miljoni sündi Prantsusmaal (James xnumx) ja 150 tuhat sündi USA-s (Ben-porath xnumx) selgus, et poisi sünnitamise tõenäosus suureneb koos vanemate vendade arvu suurenemisega ja väheneb vanemate õdede arvu suurenemisega, mis on vastuolus ESBga. Biggar jt (1999) Nende andmete põhjal viisime läbi miljoni sündi 1,4-i statistilise analüüsi ja leidsime, et vanemate vendade arvu suurenemisega kasvab poisi sündimise tõenäosus.
Neljandaks, eeldus, et perekonna esimesel sündinud poisil ei tohiks olla homoseksuaalseid eelistusi ja vastavalt sellele suureneb nende arengu oht vanemate vendade arvu suurenemisega, on pehmelt öeldes spekulatiivne.
Mitte igal homoseksuaalsel mehel pole vanemaid vendi, teisalt on mõned vanemad vennad või perekonnas ainult poisid homoseksuaalsed. Hüpoteesi toetajad esitasid vastuargumendi, mille kohaselt väidetavalt oli selliste meeste emadel enne sündi isasloote spontaanne abort, mis käivitas immuniseerimisprotsessi. Spontaansete abortidega paaride levimus on 1%; umbes pooltel neist juhtudest on lootel normaalne karüotüüp, st võib eeldada, et pooled spontaansetest abortidest on põhjustatud immuunreaktsioonidest (Alltuulekülg 2000) Spontaanse abordi tagajärjel surevate embrüote soo suhte uuringud näitavad, et enam kui pooled olid naised: meeste ja naiste suhe on 0,76 (Eiben xnumx) 0,71 (Eiben xnumx) 1,03 (Ole xnumx); 0,77 (Smith 1998) 0,77 (Evdokimova 2000) 0,83 (Morikawa xnumx) 0,35 (Halder 2006) 0,09 (Kano xnumx).
Teisest küljest tuleks immuunhüpoteesi kohaselt rünnata iga isase loote aju kõigi järgnevate raseduste ajal järjest intensiivsema intensiivsusega, st läbida üha enam ja enam “feminiseerumist”, kuid see pole nii. Kõigil gei noorematel vendadel pole homoseksuaalseid eelistusi. Huvitav on see, et soolise identiteedi rikkumisega meeste nooremad vennad - kelle aju peaksid Blanchardi hüpoteesi kohaselt läbima "feminiseerumise" - arenevad normaalselt (Roheline xnumx).

Allikas: Michael Ochsi arhiiv, Getty Images
Samuti võib hüpoteesi kohaselt eeldada, et hiljem sündinud vennad kannatavad ema suurenenud immunoloogiliste rünnakute tõttu paljude füüsiliste probleemide käes, aga vastupidi: hilisem sünnijärjestus on peamiselt seotud paranemise, mitte halvenemisega tervis (Juntunen xnumx; Cardwelli xnumx; Sorenson 2005; Richiardi xnumx).
ESB selgitav sotsiaalse mõju hüpotees
Ema immuniseerimise hüpoteesi autorid märkisid ise:
„… Muidugi on ema vennaefektile lisaks ema immuunvastuse hüpoteesile ka muid võimalikke seletusi. Kõige populaarsem konkureeriv hüpotees on see, et seksuaalne suhtlemine täiskasvanud meestega suurendab poisi homoseksuaalse tõmbe tekkimise tõenäosust ja et poisi võimalused selliseks suhtlemiseks suurenevad proportsionaalselt tema ja vanemate vendade arvuga ... ”(Ellis 2001).
Wellings ja kolleegid (1994, lk 204 - 206) leidsid, et poiste internaatkoolides käinud mehed teatasid oma elu jooksul suurema tõenäosusega homoseksuaalsetest kogemustest kui mehed, kes sellistes koolides ei käinud, kuid nende osakaal ei olnud erinev isikud, kes teatavad homoseksuaalsetest kogemustest hilisemas elus. ” Blanchard (Ellis 2001) viidatud avaldamisele Wellings ja kolleegid (1994) kui tõendit selle kohta, et sotsiaalne hüpotees pole asjakohane. Kuid nad tõlgendasid neid andmeid omapärasel viisil. Wellings lehel 206 pakub graafikut, mis näitab, et umbes 1,5% 7925 meestest, kes ei ole käinud internaatkoolis, teatasid viimase 5 aasta jooksul rohkem kui ühest homoseksuaalsest kontaktist ja 2% koolis käinud 412 meestest internaatkool. Ilmselt räägivad need andmed (rühmade ebaproportsionaalne suurus) tõenäolisemalt sotsiaalse hüpoteesi kasuks. Mõelge muudele sotsiaalteooriaga seotud uuringutele.
Blanchard ise märkis, et meessoost pedofiilidest oli umbes 25% homoseksuaalseid pedofiile (Blanchard 2000b). See on umbes kümme korda suurem homoseksuaalide osakaal nende meeste hulgas, kelle seksuaalsed huvid on suunatud täiskasvanud meestele. On oletatud, et meeste seas on homoseksuaalsusel ja pedofiilial ühine põhjus ja see põhjus on seksuaalsed (või peaaegu seksuaalsed) kogemused varases eas (James 2004). Selle idee kohaselt pärsivad varased homoseksuaalsed kogemused seksuaalse huvi teket vastassugupõlves täiskasvanueas. Rimafedi (1992. aasta) leidis, et noorukitel väheneb ebakindlus oma seksuaalsete eelistuste suhtes vanusega: need autorid viitavad sellele, et seksuaalne identiteet areneb noorukieas ja seda mõjutab seksuaalne kogemus.
Lisaks täheldatakse geide seas sagedamini seksuaalse vägivalla juhtumeid lapseeas kui heteroseksuaalsete meeste seas (Paul 2001; Finkelhor xnumx, 1984); meeste seksuaalse rünnaku ja seksuaalkuriteo vahel oli oluline seos (Klaasist 2001); märkimisväärselt suurem osa täiskasvanud meessoost homoseksuaalsetest isikutest, keda on teavitatud seksuaalvahekorra julgustamisest või sunnimisest kuni 19 aastani (Cunningham 1994); Võrreldes kontrollrühmaga täheldati homoseksuaalsete eelistuste kõrgemat määra noormehi, kes kannatasid lapsepõlves seksuaalse väärkohtlemise all (Johnson 1987; Finkelhor xnumx, 1984; Wyre sisse Tate xnumx; Cunningham xnumx; Klaasist 2001; Rind xnumx; Garcia xnumx; Arreola 2005; Beitchman xnumx; Jinich xnumx; Laumann xnumx; Lenderking 1997; Paul 2001; Tomeo 2001; Freund xnumx) Võib järeldada, et homoseksuaalsetel huvidel on olenemata tõmbeobjekti vanusest ühine põhjus. Blanchardi uuringud on näidanud, et SBE-d nähakse ka homoseksuaalsete ja biseksuaalsete pedofiilide seas, st sellistel isikutel on vanemad vennad (Bogaert 1997).
Lee jt (2002) üritasid välja selgitada, milline mitmest riskitegurist - laste emotsionaalne väärkohtlemine, käitumisprobleemid ja laste seksuaalne väärkohtlemine - oli seotud järgmistega: pedofiilia, ekshibitsionism, seksuaalne väärkohtlemine. Laste seksuaalne kuritarvitamine oli pedofiilia spetsiifiline riskifaktor. Muud seotud tegurid (emotsionaalne väärkohtlemine ja käitumisprobleemid) ei olnud pedofiiliaga nii tihedalt seotud. Lisaks, võttes arvesse selget korrelatsiooni mitme homoseksuaalse õe-venna olemasolu perekonnas ja verepilastust, tuleks intsesti pidada bioloogiliste seletuste võimaliku alternatiiviks. Kui üks vend (tavaliselt vanem) näitab üles homoseksuaalseid kalduvusi, on teistel vendadel oht võrgutada või vägistada, mis võib nende homoseksuaalse tegevuse fikseerida (Cameron 1995) Briti statistika kohaselt leiab 38% perekonnas aset leidnud seksuaalse vägivalla juhtudest aset venna poolt (Cawson xnumx) Teadlase sõnul Bartlett (2018), on populaarse psühholoogia teemal arutelud selle üle, kas täiskasvanu isiksus kujuneb sõltuvalt tema sünnijärjestusest, pikk lugu, kus on palju teaduskirjandust, mis hõlmab tuhandeid avaldatud teoseid (Damian xnumxa; Paulhus 2008; Lõhe xnumx) Viimase paarikümne aasta jooksul on selle teema uurimistöö rajatud arvamusele, et vendade ja õdedevaheline konkurents vanemate tähelepanu ressursi pärast viib faktini, et perekonnas sündivate laste sünnijärjestus mõjutab laste individuaalseid omadusi. Kuna lapsed kohanevad peres mitmesuguste niššide kasutamisega, on reeglina vanemad lapsed domineerivamad ja võtavad osa oma vanemlikest jõududest, hiljem on lapsed aga ekstravertsemad ja seltskondlikumad (Sulloway 1996) Tuleb märkida, et kuna erinev pere suurus ja sotsiaalmajanduslik seisund koos väikeste valimitega mõjutavad märkimisväärselt statistiliste arvutuste tulemusi, peaksid uuringud, milles on võimalik enam-vähem piisavalt uurida võrreldavaid ESBsid, sisaldama vähemalt 30 tuhat õdede-vendade võrdlust, samal ajal kui kuidas peetakse uuringuid, kus võrreldakse suhteliselt ühtlasi perekondade proove, alustades 500 perekondadest (Paulhus 2008) Ehkki väikeste valimitega uuringud näitavad ESB kohta vastuolulisi andmeid, on suurtes uuringutes (nt Rohrer xnumx, n = 20 000; Damian xnumxb, n = 377 000), sünnijärjekorra mõju individuaalsetele omadustele (Damian xnumxa) Need empiirilised andmed näitavad hästi taasesitatavat efekti, mille korral iga järgneva lapse intelligentsusnäitajad langevad umbes kümnendiku võrra standardhälbest, kui laps elab täiskasvanueas (Kristensen 2007), mis näitab selgelt, et mõju põhjus on vanemate investeeringute vähenemine, mitte bioloogilised emakasisesed protsessid. Suuremahulised uuringud näitavad ka sünnijärjekorra mõju sellistele omadustele nagu õpitulemused, rahaline edu ja suitsiidirisk (Bjørngaard 2013; Must xnumx).
Seega puudub samasooliste ligitõmbavuse bioloogilisel alusel, mida propageerib vendade sünnijärjekord, empiirilist tuge, samas kui selle vastu on palju empiirilisi tõendeid.
LGBT + suhtumise kahesus - Blanchardi liikumine
Oletame, et ESB ja emade immuniseerimine toimuvad ja põhjustavad muutusi käitumises. Sel juhul ühendab Blanchardi hüpotees homoseksuaalsuse ja transseksuaalsuse (nagu ka homoseksuaalse pedofiilia) - ja tänapäevases LGBT + liikumises on see jumalateotus. Näiteks vastavalt Ameerika Psühholoogilise Assotsiatsiooni andmetele on seksuaalne iha ja seksuaalne identiteet täiesti omavahel seotud nähtused (APA 2011 / 2014) Blanchardi hüpoteesi kohaselt on transseksualism patoloogia, mille põhjustab kas (1) homoseksuaalse külgetõmbe äärmuslik manifestatsioon, milles aju “feminiseerumine” on nii väljendunud, et see mõjutab ka seksuaalset enesemääratlust; või (2) vaimne hälve, milles seksuaalne külgetõmme pole suunatud mitte vastassoost, vaid iseendaga vastassoost pilti tehes (Blanchard nimetas viimast seisundit “autogynephilia”)14) (Blanchard 1989; Bailey 2003) Blanchard peab transseksualismi ühemõtteliselt patoloogiliseks nähtuseks. Lisaks märkis Blanchard ühes intervjuus:
"... ma ütleksin, et kui oleks võimalik alustada nullist, jättes tähelepanuta kogu homoseksuaalsuse DSM-ist väljajätmise ajaloo, on normaalne seksuaalsus kõik, mis on seotud paljunemisega15... "(Cameron 2013).
Selline julge seisukoht tekitab LGBT + - liikumise - eriti selles osas, mis tähistab "T" - rahulolematust.Wyndzen xnumx; Troadsmap; Dreger 2008; Serano 2010).
“LGBT +” aktivistid kirjutavad Blanchardist - liigutused:
„… Blanchardit tsiteerivad sageli LGBT-vastased rühmad (…) Ja miks mitte? Blanchard kasvas üles katoliiklasena, tal on väga traditsiooniline seisukoht, et igasugune seksuaalvahekord, mis ei hõlma peenist ja tupet, on ebanormaalne (...) Kui dr Blanchard oleks ükskõik milline pähkel ilma positsiooni ja autoriteedita, saaks ta kergesti diskrediteerida. Kuid see pole nii - vastupidi, ta oli parafiiliate ja seksuaalsete düsfunktsioonide eest vastutavas JSM-i komisjonis (...) Ta astub avalikult vastu LGBT-inimeste vastu ... "(Tannehill xnumx).
Teiselt poolt seab Blanchardi hüpoteesi kinnitamine kahtluse alla ühe LGBT + põhimõttelise dogma - liikumise - seksuaalse külgetõmbe mitmekesisuse normatiivsuse kontseptsiooni objekti soo järgi. Tõepoolest, sel juhul selgub homoseksuaalse külgetõmbe põhjus - PATHOLOOGILINE immuunvastus. Vastasel juhul peavad liikumise “LGBT +” aktivistid moonutama meditsiini ja bioloogia mõistmist nii, et arvutada välja immuunvastus, mis põhjustab raseduse katkemist, kehakaalu langust, vähenenud paljunemisvõimalusi, psühho-intellektuaalse seisundi muutust, mis nõuab hormonaalseid ravimeid ja kirurgilist sekkumist, aga ka pedofiilsed eelistused ja kalduvus vägivallale on tavaline võimalus.
Lisaks on väljavaated homoseksuaalsete eelistuste ennetamiseks poistel analoogselt Rhesus-vastaste immunoglobuliinide kasutamisega Rh-konfliktiga raseduse ajal. Milline osa tulevastest vanematest, isegi neile, kes on lojaalsed liikumisele „LGBT +”, keeldub teadlikult võimalusest vähendada oma poiste homoseksuaalse külgetõmbe riske? Tõepoolest, tänapäeval selgitatakse igale naisele hoolikalt abordi lubatavust ja rutiini. Kas naise õigus mõjutada loote elu laieneb ka õigusele mõjutada tema edasist seksuaalkäitumist või on olemas valimiskeeld ja kohtu alla andmine nende spetsialistide poolt, kes sellise võimaluse pakuvad?
Ühel või teisel viisil on need küsimused praegu tõenäolised.
Tõlgendusprobleemid
Sarnaselt eelmistes jagudes käsitletuga on empiiriliste uuringute tulemuste osas mõned olulised sisemised piirangud. Nende piirangute eiramine on üks peamisi põhjuseid, miks avalikus ruumis tehtud teadusuuringuid valesti tõlgendatakse. Nagu aju struktuuri näide näitas, on üsna ahvatlev eeldada, et kui konkreetne bioloogiline profiil on seotud mõne käitumis- või psühholoogilise tunnusega, siis on selline bioloogiline profiil selle tunnuse põhjustaja. See arutluskäik põhineb eksimisel.
Järgneva hüpoteetilise näite abil illustreerime lühidalt mõnda selle valdkonna loomupäraseid piiranguid. Oletame, et peame läbi viima joogaõpetajate ja kulturistide aju võrdleva uuringu. Kui otsite piisavalt kaua, ilmnevad lõpuks nende rühmade vahel statistiliselt olulised erinevused morfoloogilise struktuuri või aju funktsioonide mis tahes piirkonnas. Kuid see ei tähendaks, et sellised erinevused määravad joogaõpetaja ja kulturismi elutrajektooride omadused. Ajuomadused võivad olla pigem eripäraste käitumisharjumuste ja huvide tagajärg kui põhjus. Neuroplastilisuse uuringud näitavad, et vaatamata kriitilistele arenguperioodidele, mille jooksul aju muutub kiiremini ja tugevamini (näiteks väikelaste keelelise arengu ajal), jätkub aju muutumine kogu elu vältel, reageerides käitumisharjumustele (näiteks žongleerimine või mängimine pillimäng), elukogemus, psühhoteraapia, ravimid, psühholoogiline trauma ja suhted. Kasuliku ja juurdepääsetava ülevaate neuroplastilisuse uuringutest leiate Doidge 2007-ist.
Selle kindlakstegemine, kas mingil põhjusel on bioloogiline põhjus, on äärmiselt keeruline protsess ja konkreetse geneetilise seose tuvastamine on veelgi keerulisem ülesanne. Uuringud, mis pakuvad deklaratiivselt vaieldamatuid tõendeid selle kohta, et homoseksuaalid on sündinud sel viisil, on parimal juhul ebajärjekindlad ja nende tulemused on oma olemuselt suuresti korrelatsioonis.
Mõnel juhul, näiteks kaksikute uuringutes, viitavad tõendid, et varajastel keskkonnateguritel on domineeriv mõju homoseksuaalsete kalduvuste ilmnemisele. Kahe teguri omavaheline seos ei tähenda sugugi, et nende vahel oleks põhjuslik seos. Korvpallurid on pikad - korvpalli mängimine on kindlasti korrelatsioonis kiire kasvuga. “Korvpalligeeni” aga pole. Ilmselt on mõned huvitavad seosed esitatud väidetavalt põhjuslike teguritena poliitilistel ja propaganda eesmärkidel.
Lõpuks oletame, et mõnel inimesel võib olla eelsoodumus homoseksuaalsete kalduvuste tekkeks geneetilise, sünnieelse, hormonaalse mõju või muude füüsiliste või ajuomaduste tõttu. Kas see tähendab, et homoseksuaalsus on kaasasündinud nähtus? Üldse mitte selle mõistmisel, kuidas seda meedia ja populaarkultuur esindavad. Häbelikud ja kunstilised noored poisid, kelle isa ei pööranud kasvatamisele tähelepanu ega olnud õige meheliku käitumise näide, võivad olla homoseksuaalsete kalduvuste tekke ohus. Selle põhjuseks ei ole homoseksuaalne “geen”, vaid seksuaalse identiteedi kujunemise häiritud vaimse protsessi tõttu. Sellistel poistel on emotsionaalne vajadus enesejaatuse ja meeste tähelepanu järele. Sarnast pilti täheldatakse tüdrukutel, kes ei vasta klassikalistele seksuaalprofiilidele. Selliste laste probleeme ja emotsionaalseid vajadusi mängivad sageli seksuaalse ja seksuaalse maailmapildi praegused suundumused.
Need näited illustreerivad ühte levinumat probleemi, mis selliste uuringute laialdase tõlgendamise korral ilmneb - eeldus, et neurobioloogilised tegurid määravad konkreetse käitumismudeli.
Kui loodus varjab kedagi samasoolise külgetõmbega, siis miks ei anna ta sellele füüsilisi omadusi, mis on vajalikud selle realiseerimiseks? Näiteks tihe ja mitmekihiline pärasoole epiteeli membraan, mis on võimeline vastu pidama pikaajalisele hõõrdumisele, näärmetega, mis vabastavad ohtralt määrimist, peenem peenis pärasoole tungimiseks jne. Kui need omadused oleksid homoseksuaalide seas olemas, siis võiks rääkida kaasasündinud olemusest. Kui normaalse kromosoomikomplekti ja normaalse reproduktiivsüsteemi korral meelitatakse neid objekti juurde, mille abil pole seda võimalik sihtotstarbeliselt kasutada, siis tundub jutt selle nähtuse bioloogilisest seisundist väga spekulatiivne.
Mõne LGBT + liikumise esindaja arvamus
Ameerika psühholoogiline ühing avaldas 2014-is psühholoogiliste haiguste ja seksoloogia juhendi. Siin on otsesed tsitaadid sellest:
"... Praegu pole tuvastatud ühtegi geeni, mida võiks seostada homoseksuaalsusega ..." (Rosario aastal APA 2014, lk 579)
"... Vaieldamatu reaalsus on see, et inimese seksuaalkäitumise määravad paljude tegurite kombinatsioon: bioloogiline, sotsiaalne ja valitud tegur ..." (Kleinplatz APA 2014, lk 256).

APA juhtkonna mitme peatüki autor on APA ekspertkomisjoni liige professor Lisa Diamond, kes ei varja oma homoseksuaalseid eelistusi. Teemant vastandub homoseksuaalsuse geneetilise konditsioneerimise teooriale. Ta on kindel, et tees “homoseksuaalid sündisid niimoodi ja ei saa muutuda” on ekslik. 2013 aastal väitis Diamond Cornelli ülikoolis peetud loengus:
"... Ma usun, et queer-kogukond peaks lõpetama ütlemise" me oleme sündinud selliselt ja me ei saa muutuda "ja kasutada seda loosungit oma võitluses ... Ma arvan, et me ei vaja seda argumenti enam ja on isegi valus, sest tänaseks on kogunenud veenev köide teaduslikud andmed, mida teavad nii “teine pool” kui ka meie ... ”(Diamond 2013).

Paljude kunsti- ja filosoofiaraamatute autor, ameeriklane Camilla Paglia, kes ei varja oma samasooliste eelistusi, nendib otsekui:
“... homoseksuaalsus pole norm. Vastupidi, see on väljakutse normile ... Queeri teoreetikud - see kokkutõmbunud kamp freeloader-petjaid - on püüdnud minna poststrukturalistlikule kursile, väites, et normi pole, kuna kõik on juhuslik ja suhteline. See on see rumal ummiktee, kus sõnade kinnisideeks langenud inimesed langevad, kui nad on kurdid, rumalad ja pimedad ümbritseva maailma suhtes. Loodus on olemas, meeldib see teadlastele või mitte, kuid looduses on sigimine ainus ja alistamatu reegel. See on norm. Sugude kehad on loodud paljunemiseks. Peenis sobib tuppe ja ükski veider žongleerimine sõnadega ei muuda seda bioloogilist fakti ... Keegi pole sündinud homoseksuaalsena. Idee ise on naeruväärne ... Homoseksuaalsus on kohanemine, mitte kaasasündinud omadus ... "(Paglia 1994, lehed 70 - 76).
Teist prominentset Ameerika aktivisti Cynthia Nixonit ründas LGBT + - liikumine, et avalikult avaldada arvamust, et tema samasooliste motivatsiooni veab isiklik valik, mitte bioloogia (Witchell 2012).
LGBT + ameeriklaste aktivist - liikumisajakirjanik Brandon Ambrosino teatas ka, et ta ei sündinud, vaid valis teadlikult homoseksuaalse eluviisi (Ambrosino 2014), mis kutsus esile osa tema kolleege nördimusest LGBT + liikumisest (Arana xnumx).

Allikas: Frazer Harrison / WireImage
Feminist ja LGBT + aktivist - Karl Mantilla Liikumine kirjutab oma artiklis:
"... Olen juba ammu mõelnud, et" LGBT + "strateegia - liikumus kasutada argumenti sünnipärasuse kohta on uskumatult labane ... Muidugi, see on valik - kuidas saaks teisiti? ... Mõnda aega käisin naiste tugirühmas, kes otsustasid traditsioonilises abielus lesbid saada. Mingil hetkel esitasin küsimuse: "Kuidas saite aru, et olete lesbid?" Üks naine vastas, et ta ei tundnud end kunagi meestega emotsionaalselt lähedal ja et naised mõistavad teda alati paremini. Teine ütles kohe, et ka tema tundis, et suudab emotsionaalselt avatud olla ainult naistega. Teised noogutasid nõusolevalt. Mis selles olukorras valesti oli? Peaaegu kõik naised tunnevad seda! Iga heteroseksuaalne naine, keda ma olen kunagi tundnud, tundis end oma sõpru usaldades mugavamalt, tundis end neile lähemal, tundis end paremini mõistetuna ja avas naisi. Kui selleks on vaja lesbi saada, siis on kõik naised lesbid. See on sama vana kui maailm ... naiste kaebused, et nende mehed ei räägi nendega, ei mõista nende tundeid ega huvita, mida nad ütlevad. Naisteajakirjade kõige levinumad artiklid on see, kuidas panna oma mees ennast avama ja teiega rääkima ... emotsionaalse läheduse tundel inimesega pole mingit bioloogilist alust, see on tingitud inimese emotsionaalsetest ja psühholoogilistest omadustest ... aja jooksul sai mulle selgeks, et naised see tugirühm tundis lihtsalt tohutut süütunnet oma abikaasa lahkumise pärast ... Nii et mõte, et nad ei saa midagi teha selle vastu, et nad on lesbid, et sellel on bioloogiline põhjus, vabastas nad süüst ja vastutusest oma tegude eest ... "(Mantilla xnumx).
LGBT + aktivist, Californias asuv liikumine nimega Gail Madwin, on isegi loonud terve saidi, kus väidetakse, et homoseksuaalne käitumine pole kaasasündinud, vaid teadliku valiku tõttu (Queer by choice). LGBT + endine aktivist, David Benkofi liikumine põhjendab ka seda, et homoseksuaalset elustiili ei määra mingil juhul mingid bioloogilised tegurid (Benkof xnumx).
Märkused
1: me sündisime niimoodi
2 ei ole üldiselt üksteisega seotud
3 homoseksuaalsete kalduvuste "range" kriteeriumi järgi: 2 ja rohkem nn Kinsey skaala.
Inglise keeles 4 GWAS, genoomsete ühenduste uuringud
5 võttis teadusringkondades kasutusele konverentsidel CV-de esitamise tava - lühikese artikli, tavaliselt 150 - 250 sõnade suurus -, millele järgneb terve artikli avaldamine ajakirjas
6 English: tõenäoliselt sündinud eelsoodumusega
7 Sellega seoses võib tulemuste levitamine inimese kohta olla piiratud
8 viriliseerumine - meditsiiniline termin rikkumiseks, mille käigus naiste seksuaalsed omadused muutuvad meessoost
9 English: “eesmise hüpotalamuse (INAH) interstitsiaalsed tuumad”
Eesti keeles 10: inimese jahmunud reaktsiooni (impulsi pärssimine enne impulsi) (PPI)
11 English: “vennaliku sünnijärjekorra efekt (FBO)”
12 Vaadake Twin Research sektsiooni
13 Lisaks on antigeenid PK ja siiriku hülgamisreaktsioonide korral individuaalsed (PK korral isalised), kuid isastele omased.
Kreeka keelest 14 autos - “ise”, gini - “naine” ja filia - “armastus”; "Armastus naise vastu"
15 Ma ütleksin, et kui alustada nullist, ignoreerida kogu homoseksuaalsuse eemaldamise ajalugu DSM-ist, on normaalne seksuaalsus ükskõik, mis on seotud paljunemisega
lisainfo
Lisateavet ja üksikasju leiate järgmistest allikatest:
1. Whitehead NE, Whitehead BK. Minu geenid panid mind seda tegema! Homoseksuaalsus ja teaduslikud tõendid. Whitehead Associates. Väljaanne 5th 2018.
2. Mayer LS, McHugh PR. Seksuaalsus ja sugu: järeldused bioloogilistest, psühholoogilistest ja sotsiaalteadustest. Uus Atlantis, Number 50, Sügisel 2016.
3. Sprigg P., et al. Täpsemalt öeldes: mida uuringud näitavad homoseksuaalsuse kohta. Washington: Perekonnauuringute nõukogu (2004).
3. Harrub B, Thompson B, Miller D. “See on viis, mille Jumal mind tegi” - Homoseksuaalsuse ja “geeni geeni” teaduslik uurimine. Põhjus ja ilmutus. August 2004; 24 (8): 73.
5. Sorba r. “Sündinud gei” petmine. Ryan Sorba Inc. Esimene väljaanne 2007.
6. Valgepea NE. Poistevastane antikeha? Ema immuunsuse hüpoteesi uuesti läbivaatamine. Ajakiri Biosocial Science 2007.
7. Rüütel r. Sündinud või aretatud? Teadus ei toeta väidet, et homoseksuaalsus on geneetiline... Kultuuri- ja pereinstituut. Naised Ameerika pärast. 2004.
8. van den Aardweg G. Homoseksuaalsus ja bioloogilised tegurid: tõelised tõendid - puuduvad; Eksitavad tõlgendused: palju. Kordustrükk NARTH bülletäänist, talv 2005.
9. Hubbard R, Wald E Geenimüüdi plahvatamine: kuidas teadlased, arstid, tööandjad, kindlustusettevõtted, koolitajad ja korrakaitsjad geneetilisi andmeid loovad ja manipuleerivad. Beacon Press, Boston; 1999.
Bibliograafilised allikad
- Vassilchenko G.S. Seksopatoloogia: käsiraamat / toim. G.S. Vassilchenko. - M., 1990.
- Yarygin V.N. (2003) // Bioloogia. 2-i raamatus Toim. V.N. Yarygin / Yarygin V. N., Vasilieva V. I., Volkov I. N., Sinelshchikova V. V. 5 ed., Rev. ja lisage. - M.: Kõrgkool, 2003. Raamat 1 - 432., Raamat 2 - 334.
- ASHG 2015. Epigeneetiline algoritm ennustab täpselt meeste seksuaalse orientatsiooni tulemusi, millest teatati ASHG 2015 aastakoosolekul. Koheseks vabastamiseks neljapäeval, oktoobril 8, 2015 http://www.ashg.org/press/201510-sexual-orientation.html
- Albrecht ED, Pepe GJ. Platsenta angiogeneesi ja loote munasarjade arengu östrogeenregulatsioon primaatide raseduse ajal ”, Rahvusvaheline Arengubioloogia Ajakiri 54, nr. 2 - 3 (2010): 397 - 408, http://dx.doi.org/10.1387/ijdb.082758ea
- Allen S. „Geenigeeni” probleemne jaht. Igapäevane metsaline. 20.11.2014. https://www.thedailybeast.com/the-problematic-hunt-for-a-gay-gene (01.12.2017 on kinnitatud)
- Ambrosino B. Ma ei olnud nii sündinud. Valin olla gei. Uus Vabariik. 28. jaanuar 2014. https://newrepublic.com/article/116378/macklemores-same-love-sends-wrong-message-about-being-gay
- APA Ameerika psühholoogiline ühendus. Vastused küsimustele. Transsooliste inimeste, sooväljenduse ja soolise identiteedi kohta. Koostatud avalike suhete büroo ja ühingu liikmete poolt. Trükitud 2011; värskendatud 04 / 2014.https: //www.apa.org/topics/lgbt/transgender-russian.pdf
- Arana G. Ezra Kleini Queeri uus palk. 13. märts 2014. Ameerika väljavaade.
- Arreola, SG, Neilands, TB, Pollack, LM, Paul, JP & Catania, JA (2005) Laste seksuaalse väärkohtlemise suurem levimus meestega seksivate latino meeste seas kui meestega seksivad mitte-latino mehed: andmed Linnameeste terviseuuring. Laste väärkohtlemine ja hooletusse jätmine 29, 285–290.
- Bailey J. M, et al., “Inimese meeste homoseksuaalsuse emade stressiteooria test”, seksuaalkäitumise arhiiv 20, nr. 3 (1991): 277 - 293, http://dx.doi.org/10.1007/BF01541847
- Bailey, J. Michael (2003). Mees, kes oleks kuninganna: soolise paindumise ja transseksuaalsuse teadus. Joseph Henry Press
- Bailey JM jt. Geneetilised ja keskkonnamõjud seksuaalsele orientatsioonile ja selle korrelatsioonid Austraalia kaksikproovis. J Pers Soc Psychol. 2000 Mar; 78 (3): 524-36.
- Bains JS, Wamsteeker Cusulin JI, Inoue W. Stressist tingitud sünaptiline plastilisus hüpotalamuses. Nat Rev Neurosci. 2015 juuli; 16 (7): 377-88. doi: http://dx.doi.org/10.1038/nrn3881
- Parun M. Geneetika ja inimese seksuaalne sättumus. Bioloogiline psühhiaatria. Juuni 1 - 15, 1993, köide 33, annab välja 11-12, lehed 759 - 761.
- Bartlett NT, Hurd PL.Emaise sünnikorralduse mõju isiksusele: kas mõistlikud nõuded nõuavad erakorralisi tõendeid? Arch Sex Behav. 2018 jaan; 47 (1): 21-25. doi: 10.1007 / s10508-017-1109-z.
- Be, G., Velasquez, P. & Youlton, R. (1997) Spontaanne abort: tsütogeneetiline uuring 609 juhtumist. Revista Medica de Chile 125, 317-322.
- Bearman PS, Brückner H. Vastand - seks kaksikud ja noorukid sama - seksi atraktsioon. Ameerika sotsioloogiaajakiri 2002 107: 5, 1179-1205
- Karu, PS, ja Brückner, H. (2002). Vastassugupoole kaksikud ja teismeliste samasooliste külgetõmme. American Journal of Sociology, 107, 1179–1205. doi: 10.1086 / 341906.
- Beer, AE ja Billingham, RE (1975) Piima immunoloogilised eelised ja ohud ema-perinataalses suhtes. Sisehaiguste arhiivid 83, 865-871.
- Beitchman, JH, Zucker, KJ, Hood, JE, Da Costa, GA & Akman, S. (1991) Laste seksuaalse väärkohtlemise lühiajaliste mõjude ülevaade. Laste väärkohtlemine ja hooletusse jätmine 15, 537–556.
- Benkof D. Keegi ei ole nii sündinud, ütlevad geiajaloolased. Igapäevane helistaja. 19.03.2014. dailycaller.com/2014/03/19/nobody-is-born-that-way-gay-historians-say/
- Ben-Porath, Y., et al. (1976) Kas seksieelistused on tõesti olulised? QJ Econ. 90, 285 - 307.
- Berenbaum SA. Kuidas mõjutavad hormoonid käitumise ja närvide arengut: sissejuhatus eriväljaandesse „Gonadaalsed hormoonid ja seksuaalsed erinevused käitumises. Arengu neuropsühholoogia 14 (1998): 175 - 196, http://dx.doi.org/10.1080/87565649809540708
- Biggar, RJ, et al. (1999) Suhtarvud, perekonna suurus ja sünnijärjestus. Olen J. Epidemiol. 150, 957 - 962.
- Arved, Beckwith. Tehnoloogia ülevaade. Juuli 1993, lk 60.
- Bjørngaard, JH, Bjerkeset, O., Vatten, L., Janszky, I., Gunnell, D., & Romundstad, P. (2013). Emapoolne vanus lapse sünnil, sündimus ja enesetapp noores eas: A vendade võrdlus. American Journal of Epidemiology, 177, 638-644. https://doi.org/10.1093/aje/kwt014.
- Must, SE, Devereux, PJ ja Salvanes, KG (2005) Mida rohkem, seda uhkem? Pere suuruse ja sünnikorra mõju laste haridusele. Quarterly Journal of Economics, 120, 669–700. https://doi.org/10.2307/25 098749.
- Blanchard R (august 1989). "Mittehomoseksuaalsete sooliste düsfooriate klassifitseerimine ja märgistamine." Seksuaalkäitumise arhiiv. 18 (4): 315–34. doi:10.1007/BF01541951
- Blanchard R, Bogaert AF. (1996) Meeste homoseksuaalsus ja vanemate vendade arv. American Journal of Psychiatry 153, 27 - 31.
- Blanchard R, Bogaert AF. Meeste homoseksuaalsus ja vanemate vendade arv. American Journal of Psychiatry; Jan 1996a; 153, 1; Teadusraamatukogu, lk. Xnumx
- Blanchard R., et al. (2000) Vendade sünnijärjestus ja seksuaalne sättumus pedofiilides. Archs seksuaalse käitumise 29, 463 - 478.
- Blanchard, R. & Bogaert, AF (1996b) Homoseksuaalsete ja heteroseksuaalsete meeste biodemograafiline võrdlus Kinsey intervjuu andmetes. Seksuaalkäitumise arhiivid 25, 551–579.
- Blanchard, R. & Bogaert, AF (1998) Laste, puberteediealiste ja täiskasvanute homoseksuaalsete ja heteroseksuaalsete kurjategijate sünnijärjekord. Seksuaalkäitumise arhiivid 27, 595-603.
- Blanchard, R. & Ellis, L. (2001) Sünnikaal, seksuaalne orientatsioon ja eelnevate õdede-vendade sugu. J. biosots. Sci. 33, 451-467.
- Blanchard, R. (2014). Perekondlike suuruste erinevuste tuvastamine ja parandamine seksuaalse sättumuse ja vennaliku sünnijärjekorra uurimisel.Seksuaalse käitumise arhiivid, 43, 845 - 852. https://doi.org/10.1007/s10508-013-0245- 3.
- Blanchard, R. Vennalik sünnijärjestus, perekonna suurus ja meeste homoseksuaalsus: 25 aastat hõlmavate uuringute meta-analüüs. Arch Sex Behav (2018) 47: 1. https://doi.org/10.1007/s10508-017-1007-4
- Blanchard, R., & Bogaert, AF (2004). Homoseksuaalsete meeste osakaal, kes võlgnevad oma seksuaalse sättumuse vennalikule sündimisjärjekorrale: prognoosige rahvusvaheliste tõenäosuse proovide põhjal. American Journal of Human Biology, 16, 151–157.
- Blanchard, R., & VanderLaan, DP (2015). Kommentaar Kishidale ja Rahmanile (2015), sealhulgas meeste vendliku sünnijärjekorra ja seksuaalse sättumuse asjakohaste uuringute metaanalüüs. Seksuaalse käitumise arhiivid, 44, 1503-1509. doi: 10.1007 / s10508-015-0555-8
- Blanchard, R., Barbaree, HE, Bogaert, AF, Dickey, R., Klassen, P., Kuban, ME & Zucker, KJ (2000) Vennalik sünnikord ja seksuaalne orientatsioon pedofiilides. Seksuaalkäitumise arhiivid 29, 463–478.
- N-lahter: “Kuidas võib ärrituvus eksitada rassi suhtes”, tunnetus 56, nr. 2 (1995): 103 - 104, http://dx.doi.org/10.1016/0010-0277(95)00678-R
- Bogaert, AF (2003). Vanemate vendade koosmõju ja sootüübid meeste seksuaalse sättumuse ennustamisel. Seksuaalse käitumise arhiivid, 32, 129 - 134.
- Bogaert, AF (2004) .Meeste homoseksuaalsuse levimus: vennaliku sünnijärjekorra ja perekonna suuruse erinevuste mõju. Teoreetilise bioloogia ajakiri, 230, 33 - 37.
- Bogaert, AF (2005). Sooline roll / identiteet ja õdede-vendade sugu suhe homoseksuaalides. JournalofSexandMaritalTherapy, 31,217 - 227. https: // doi. org / 10.1080 / 00926230590513438.
- Bogaert, AF (2006) Bioloogilised versus mittebioloogilised vanemad vennad ja meeste seksuaalne sättumus. Riikliku Teaduste Akadeemia toimetised 103, 10771 - 10774.
- Bogaert, AF, Bezeau, S., Kuban, M. & Blanchard, R. (1997) Pedofiilia, seksuaalne orientatsioon ja sündimus. Ebanormaalse psühholoogia ajakiri 106, 331-335.
- Bogaert, AF, & Skorska, M. (2011). Seksuaalne orientatsioon, vennalik sünnijärjekord ja ema immuunsuse hüpotees: Areview. Piirid neuroendokrinoloogias, 32, 247-254.
- Bogaert, AF (2005) .Soolise soo suhe ja seksuaalne sättumus meestel ja naistel: uued testid kahes riiklikus tõenäosusproovis.Seksuaalse käitumise arhiivid, 34, 111 - 116. doi: 10.1007 / s10508-005-1005-9.
- Bogaert, AF (2010) .Füüsikaline areng ja seksuaalne orientatsioon meestele ja naistele: NATSAL-2000 analüüs. Seksuaalse käitumise arhiivid, 39, 110 - 116.doi: 10.1007 / s10508-008-9398-x.
- Briggs WM. Väidetavalt äsja avastatud “geenigeenidel” või modellioskuse olulisusel. Oktoober 13, 2015. wmbriggs.com/post/17053/
- Byne W, Tobet S, Mattiace LA jt. Inimese eesmise hüpotalamuse interstitsiaalsed tuumad: soo, seksuaalse sättumuse ja HIV staatuse erinevuste uurimine. Horm Behav. 2001 september; 40 (2): 86-92. http://dx.doi.org/10.1006/hbeh.2001.1680
- Byne W. Bioloogilised tõendid on väljakutse saanud. Teadus-Ameerika, mai 1994, lk 50 - 55.
- Caldwell, JC (1997). Stabiilse globaalse rahvastikuni jõudmine: mida oleme õppinud ja mida peame tegema. Tervise ülemineku ülevaade, 7, 37 - 42.
- Cameron P, et al. Kas verepilastus põhjustab homoseksuaalsust? Psühholoogilised aruanded, 1995, 76, 611-621.
- Cameron L. Kuidas psühhiaater, kes kirjutas seksikäsiraamatu, rääkis seksist? Emaplaat. Aprill 11 2013. https://motherboard.vice.com/en_us/article/ypp93m/heres-how-the-guy-who-wrote-the-manual-on-sex-talks-about-sex
- Cantor, JM, Blanchard, R., Paterson, AD & Bogaert, AF (2002) Kui paljud homoseksuaalsed mehed võlgnevad oma seksuaalse sättumuse vennaliku sünnijärjekorra eest? Seksuaalkäitumise arhiivid 31, 63–71.
- Cardwell, CR, Carson, DJ & Patterson, CC (2005) Vanemate vanus sünnituse ajal, sünnijärjekord, sünnikaal ja rasedusaeg on seotud 1. tüüpi diabeedi riskiga lapsepõlves: Suurbritannia piirkondlik retrospektiivne kohordiuuring. Diabeetiline meditsiin 22 200-206.
- Cawson P, et al. Laste väärkohtlemine Ühendkuningriigis: väärkohtlemise ja tähelepanuta jätmise uuring. NSPCC uurimistulemused november 2000.
- Chaix, R., Cao, C., & Donnelly, P. (2008). Kas inimesel on kaaslase valik MHC-st sõltuv? PLoS Genetics, 4, e1000184.
- Cohen-Kettenis PT, soo muutus 46, XY inimestel, kellel on 5α-reduktaasi-2 defitsiit ja 17β-hüdroksüsteroiddehüdrogenaasi-3 defitsiit. Seksuaalse käitumise arhiiv 34, nr. 4 (2005): 399 - 410, http://dx.doi.org/10.1007/s10508-005-4339-4
- Collins FS Jumala keel. New York, NY Simon & Schuster, Inc. 2006.
- Cote, K., Earls, CM & Lalumiere, ML (2002) Sünnikorraldus, sünnide intervall ja hälbivad seksuaalsed eelistused seksuaalkurjategijate seas. Seksuaalne väärkohtlemine: ajakiri Research and Treatment 14, 67–81.
- Cunningham, RN, et al. (1994) Füüsilise ja seksuaalse kuritarvitamise seostamine HIVi riskikäitumisega noorukieas ja noores täiskasvanueas: mõju rahva tervisele. Laste väärkohtlemine Negl. 18, 233 - 245.
- Damian, RI ja Roberts, BW (2015a). Arutelu seadmine sünnijärjekorra ja isiksuse üle. Rahvusliku Teaduste Akadeemia toimetised, 112, 14119-14120. https://doi.org/10.1073/pnas.1519064112.
- Damian, RI ja Roberts, BW (2015b). Sünnikorra seos isiksuse ja intelligentsusega USA keskkooliõpilaste esinduslikus valimis. JournalofResearchinPersonality, 58,96-105. Https: // doi.org/10.1016/j.jrp .2015.05.005.
- Dankers, MK, Roelen, D., Korfage, N., de Lange, P., Witvliet, M., Sandkuiil, I., Doxiadis, II & Claas, FH (2003) Isaste HLA I klassi antigeenide diferentsiaalne immunogeensus rasedatel naised. Inimese immunoloogia 64, 600-606.
- Davis N. Looduslikud sündinud tapjad: mõrvale kalduvad inimesed, näitavad uuringud. The Guardian. 28.09.2016. https://www.theguardian.com/science/2016/sep/28/natural-born-killers-humans-predisposed-to-study-suggests (01.12.2017 Verified)
- Dawkins R. Kuradi kaplan: mõtisklusi lootuse, valede, teaduse ja armastuse üle. Esimene Mariner Books'i väljaanne 2004
- Teemant Lisa. Kui erinevad on naiste ja meeste seksuaalne orientatsioon? 17.10.2013. Cornelli ülikool. https://www.youtube.com/watch?v=m2rTHDOuUBw&feature=youtu.be&t=43m13s (otsitud 01.12.2017)
- Doidge Norman, aju, mis ennast muudab: lood isikliku võidukäiguga ajuteaduse piiridest (New York: Penguin, 2007)
- Dörner Günter jt, “Stressiüritused kahe- ja homoseksuaalsete meeste sünnieelses elus”, eksperimentaalne ja kliiniline endokrinoloogia 81, nr. 1 (1983): 83 - 87, http://dx.doi.org/10.1055/s-0029-1210210
- Drabant EM jt, „Seksuaalse orientatsiooni uurimine kogu genoomi ulatuses suures veebipõhises kohordis”, 23andMe, Inc. (Programmi number: 2100W) Esitati Ameerika inimgeneetika seltsi 62. aastakoosolekul 7. novembril 2012 Californias San Franciscos. http://abstracts.ashg.org/cgi-bin/2012/ashg12s?author=drabant&sort=ptimes&sbutton=Detail&absno=120123120&sid=320078
- Dreger AD. Vaidlus mehega, kes oleks kuninganna: teaduse, identiteedi ja seksi poliitika juhtumiajalugu Interneti-ajastul. Seksuaalse käitumise arhiivid. 2008; 37 (3): 366-421. doi: 10.1007 / s10508-007-9301-1.
- Ebstein Richard P. jt, “Inimese sotsiaalse käitumise geneetika”, Neuron 65, nr. 6 (2010): 831– 844, http://dx.doi.org/10.1016/j.neuron.2010.02.020
- Eiben, B., Bahr-Porsch, S., Borgman, S., Gatz, G., Gellert, G. & Goebel, R. (1990) 750 spontaanse abordi tsütogeneetiline analüüs koorionvillide otsese ettevalmistamise meetodiga ja selle mõju raseduse raiskamise geneetiliste põhjuste uurimisele. American Journal of Human Genetics 47, 656-663.
- Eiben, B., Borgman, S., Schubbe, I. & Hansman, I. (1987) 140 spontaanse abordija koorionvillide tsütogeenne uuringusuund. Inimgeneetika 77, 137-141.
- Ellis L, Blanchard R (2001) Sünnijärjestus, õdede ja vendade sugu suhe ning raseduse katkemine homoseksuaalsete ja heteroseksuaalsete meeste ja naiste seas. Isiklik indiviid. Erinevused 30, 543 - 552.
- Ellis Lee ja Cole-Harding Shirley, „Sünnieelse stressi ning sünnieelse alkoholi ja nikotiini kokkupuute mõju inimese seksuaalsele sättumusele“, Physiology & Behavior 74, nr. 1 (2001): 213-226, http://dx.doi.org/10.1016/S0031-9384(01)00564-9
- Ellis Lee jt, „Raseduse ajal võib ema tugev stress avaldada mõju inimese järglaste seksuaalsele orientatsioonile,” Journal of Sex Research 25, nr. 2 (1988): 152 - 157, http://dx.doi.org/10.1080/00224498809551449
- Ennis D. Inimõiguste kampaania tutvustab Johns Hopkinsi vaatamisväärsusi pärast vastuolulist transraportit. 2016. NBC uudised.
- Evdokimova, VN, Nikitina, TV, Lebedev, IN, Sulchanova, NN & Nazarenko, SA (2000) Sooline suhe inimese varases embrüonaalses suremuses. Ontogenez 31, 251-257.
- Fausto-Sterling A., Balaban E. Geneetika ja meeste seksuaalne orientatsioon. Teadus. 1993; 261: 1257. http://dx.doi.org/10.1126/science.8362239
- Finkelhor, D. (1979) Seksuaalselt ohvriks langenud lapsed. Free Press, New York.
- Finkelhor, D. (1984) Laste seksuaalne väärkohtlemine: uus teooria ja uurimistöö. Free Press, New York.
- Finn R. Seksuaalsuse kuumutamise bioloogiline määramine uurimisvaldkonnana. Teadlane 10 [1]: jaan. 08, 1996.
- Flannery, KA & Liderman, J. (1994) Immunoreaktiivse teooria test immuunhäirega naiste järglaste neurodevelopmental-häirete tekkeks. Cortex 30, 635-645
- Francis AM (2008). Perekondlik ja seksuaalne sättumus: meeste ja naiste homoseksuaalsuse perekondlikud demograafilised korrelatsioonid. Seksusuuringute ajakiri, 45, 371 - 377. doi: 10.1080 / 00224490802398357.
- Freund, K. & Kuban, M. (1994) Pedofiilia väärkohtlejate teooria alus: varasema uuringu edasiarendus. Seksuaalkäitumise arhiivid 23, 553–563.
- Frisch, M., ja Hviid, A. (2006). Heteroseksuaalsete ja homoseksuaalsete abielude lapsepõlvekorrelatsioon: kahe miljoni taanlase riiklik kohordiuuring. Seksuaalse käitumise arhiiv, 35,533 547-10.1007. Doi: 10508006 / s9062-2-XNUMX.
- Garcia, J., Adams, J., Friedman, L. & East, P. (2002) Seosed mineviku väärkohtlemise, enesetapumõtete ja seksuaalse sättumuse vahel San Diego kolledži üliõpilaste seas. American College Healthi ajakiri 51, 9–14.
- Gasparoni, A., Avanzini, A., Ravagni Probizer, F., Chirico, G., Rondini, G. & Severi, F. (1992) IgG alaklassid võrreldes ema ja nabanööri seerumi ja rinnapiimaga. Lapsepõlves haiguste arhiiv 67 (1), erinumber, 41–43.
- Gavrilets S, Friberg U, Rice WR. Homoseksuaalsuse mõistmine: liikumine mustrilt mehhanismidele. Arch Sex Behav. 2017. DOI 10.1007 / s10508-017-1092-4
- Gelman M. Gay geeni tabloidi hüpe värskendus. Statistiline modelleerimine, põhjuslikud järeldused ja ühiskonnaõpetus. Oktoober 10, 2015. https://andrewgelman.com/2015/10/10/gay-gene-tabloid-hype-update/
- Ginalksi, K., Rychlewski, L., Baker, D. & Grishin, NV (2004) Valgu struktuuri ennustamine inimese Y-kromosoomi meesspetsiifilise piirkonna jaoks. Rahvusliku Teaduste Akadeemia toimetised 101, 2305-2310
- Glasser, M., et al. (2001) Laste seksuaalse väärkohtlemise tsükkel: seosed ohvriks olemise ja vägivallatsejaks saamise vahel. Br. J. Psychiat. 179, 482 - 494.
- Gomes AR, Souteiro P, SIlva CG jt. Testosterooni puudulikkuse levimus HIV-nakatunud meestel retroviirusevastase ravi ajal. BMC nakatada Dis. 2016; 16: 628. Avaldatud veebis 2016 november 3. http://dx.doi.org/10.1186/s12879-016-1892-5
- Greally J. Nädala ületõlgendatud epigeneetika uuring (2). EpgntxEinstein. New Yorgi Bronxis asuva Albert Einsteini meditsiinikolledži epigenoomikakeskuse ajaveeb.
- Green, R. (2000) Sünnijärjestus ning vendade ja õdede suhe transseksuaalides. Psühholoogiline meditsiin 30, 789 - 795.
- Gualteri, T. & Hicks, RE (1985) Meeste valikulise viletsuse immunoreaktiivne teooria. Käituma. Brain Sci. 8, 427-477.
- Guleria I, Sayegh, MH. Loote emane aktsepteerimine: tõeline inimtolerants. J Immunol märts 15, 2007, 178 (6) 3345-3351; DOI: https://doi.org/10.4049/jimmunol.178.6.3345
- Haler, A. & Fauzdar, A. (2006) Võltsitud sugude suhe ja madal aneuploidia korduva varajase vahelejäänud abordi korral. India meditsiiniliste uuringute ajakiri 124, 9–10.
- Hall Lynn S. ja Love Craig T., “Naise monosügootsete kaksikute sõrme-pikkuse vahekorrad, mis ei võimalda seksuaalset orientatsiooni”, seksuaalkäitumise arhiiv 32, nr. 1 (2003): 23 - 28, http://dx.doi.org/10.1023/A:1021837211630
- Hamer D, Copeland P. Soovide teadus: geenigeeni otsing ja käitumise bioloogia. Simon ja Schuster 1994
- Hamer D. Jumala geen: kuidas on usk meie geenidesse ühendatud. Doubleday xnumx
- Hamer DH jt, “Seos X-kromosoomi DNA-markerite ja meeste seksuaalse sättumuse vahel”, Science 261, nr. 5119 (1993): 321 - 327, http://dx.doi.org/10.1126/science.8332896
- Han, TH, Chey, MJ & Han, KS (2006) Granulotsüütide antikehad neutropeeniaga Korea vastsündinutel. Korea meditsiiniseltsi ajakiri 21, 627–632.
- Harrison Halstead, „Tehniline kommentaar paberlehele“ Seksuaalse orientatsiooniga seotud erinevused inimese käivitumise reageerimise impulsside pärssimises ”.” Washingtoni ülikooli veebisait, 15, detsember 2003, http://www.atmos.washington.edu/ ~ harrison / report / rahman.pdf.
- Hatton GI. Funktsiooniga seotud plastilisus hüpotalamuses. Annu Rev Neurosci. 1997; 20: 375-97. http://dx.doi.org/10.1146/annurev.neuro.20.1.375
- Hoekzema E, et al. Rasedus põhjustab inimese aju struktuuri pikaajalisi muutusi. Looduse neuroteaduse maht 20, lehed 287 - 296 (2017).
- Heston, L.L., Shields, J., "Homoseksuaalsus kaksikutes, perekonnauuring ja registriuuring" Arch Gen Psychiat. 1968; 18:149
- Hildebrand, H., Finkel, Y., Grahnquist, L., Lindholm, J., Ekbom, A. & Aksling, J. (2003) Laste põletikulise soolehaiguse muutuv muster Põhja-Stockholmis 1990–2001. Gut 52 1432– 1434.
- Hines M. Sünnieelne sisesekretsioon mõjutab seksuaalset sättumust ja seksuaalselt diferentseeritud lapsepõlvekäitumist. Eesmine neuroendokrinool. 2011 aprill; 32 (2): 170 - 182. doi: 10.1016 / j.yfrne.2011.02.006
- Hines Melissa jt, “Sünnieelne stress ja sooline rollikäitumine tüdrukutel ja poistel: pikisuunaline, populatsiooniuuring”, hormoonid ja käitumine 42, nr. 2 (2002): 126 - 134, http://dx.doi.org/10.1006/hbeh.2002.1814
- Hönekopp J et al., “Teise ja neljanda numbri pikkuse suhe (2D: 4D) ja täiskasvanute suguhormoonide tase: uued andmed ja metaanalüütiline ülevaade”, Psychoneuroendocrinology 32, nr. 4 (2007): 313 - 321, http://dx.doi.org/10.1016/j.psyneuen.2007.01.007
- Horgan, John. (1995) “Geigeenid, uuesti läbi vaadatud.” Scientific American, vol. 273, ei. 5, 1995, lk. 26 - 26. JSTOR, JSTOR, www.jstor.org/stable/24982058
- Hubbard R., Wald E. Geenimüüdi plahvatamine: kuidas geneetilisi andmeid loovad ja manipuleerivad teadlased, arstid, tööandjad, kindlustusettevõtted, koolitajad ja korrakaitsjad. 1999. Boston Press. ISBN: 978-080700431-9, lehel 95 - 96.
- Huffpost 2017. Dean Hamer ja Joe Wilson. https://www.huffingtonpost.com/author/qwavesjoe-855 (01.12.2017 on kinnitatud)
- Hughes IA jt, “Androgeeni tundlikkuse sündroom”, The Lancet 380, nr. 9851 (2012): 1419 - 1428, http://dx.doi.org/10.1016/S0140-6736%2812%2960071-3
- Inimese genoomi ressursid NCBI 2017-is. https://www.ncbi.nlm.nih.gov/projects/genome/guide/human/
- Izetbegovic S. ABO ja RhD sobimatus Rh-negatiivsete emadega. Materia Socio-Medica. 2013; 25 (4): 255-258. doi: 10.5455 / msm.2013.25.255-258.
- James WH. Kaks hüpoteesi meeste homoseksuaalsuse ja pedofiilia põhjustel. J.biosoc.Sci, (2006) 38, 745 - 761, doi: 10.1017 / S0021932005027173
- James, WH (1975) Suhe ja olemasolevate õdede-vendade sugukompositsioon. Ann. hum. Genet. 38, 371 - 378.
- James, WH (1985) Väidetav eelnevate vendade mõju soo suhtele. Käitumine. Aju Sci. 8, 453.
- James, WH (1996) Tõendid selle kohta, et imetajate soo suhteid sündides kontrollitakse viljastumise ajal osaliselt vanemate hormoonide taseme järgi. Teoreetilise bioloogia ajakiri 180, 271 - 286.
- James, WH (2004) Vennaliku sünnijärjekorra põhjus (ed) meeste homoseksuaalsuses. Ajakiri Biosocial Science 36, 51 - 59, 61 - 62.
- James, WH (2004b) Täiendavad tõendid selle kohta, et imetajate sooline suhe sünnil on lapseoote ajal osaliselt vanemate hormoonide taseme kontrolli all. Inimese paljunemine 19, 1250 - 1256.
- Jinich, S., Paul, JP, Stall, R., Acree, M., Kegeles, S., Hoff, C. & Coates, T. (1998) Laste seksuaalne kuritarvitamine ja HIV-i riskikäitumine homo- ja biseksuaalsete meeste seas ... AIDS ja käitumine 2, 41–51.
- Johnson, RL & Shrier, DK (1987) Meessoost patsientide varasem seksuaalne ohverdus noorukite meditsiinikliinikus. Olen. J. Psychiat. 144, 650-652.
- Juntunen, KS, Laara, EM & Kauppila, AJ (1997) Suurejooneline mitmekordsus ja sünnikaal. Sünnitusabi ja günekoloogia 90, 495-499.
- Kallmann, Franz J., “Meeste homoseksuaalsuse geneetiliste aspektide võrdlev võrdlusuuring”, Närvi- ja vaimsete haiguste ajakiri 115, nr. 4 (1952): 283 - 298
- Kano, T., Mori, T., Furudono, M., Kanda, T., Maeda, Y., Tsubokura, S., Ushiroyama, T. & Ueki, M. (2004) Abortide ja vastsündinute soolised erinevused naistel korduvate alloimmuunsete abortidega. Reproduktiivne biomeditsiin Online 9, 306-311.
- Kendler KS jt, “Seksuaalne orientatsioon USA kaksikute ja Nontwini õdede-vendade paaride rahvuslikus proovis”, American Journal of Psychiatry 157, nr. 11 (2000): 1843 - 1846, http://dx.doi.org/10.1176/appi.ajp.157.11.1843
- Kishida, M. ja Rahman, Q. (2015). Vennalik sünnijärjekord ja äärmuslik parempoolsus meeste seksuaalse sättumuse ja soolise mittevastavuse ennustajatena. Seksuaalkäitumise arhiivid, 44, 1493-1501. https: // doi. org / 10.1007 / s10508-014-0474-0.
- Kleinplatz & Diamond 2014, APA käsiraamat, 1. köide, lk 256–257
- Kolb B, Whishaw IQ. Aju plastilisus ja käitumine. Psühholoogia aastaülevaade. Vol. 49: 43-64. https://doi.org/10.1146/annurev.psych.49.1.43
- Kranz F jt, „Näo tajumist moduleerib seksuaalne eelistus“, praegune bioloogia 16, nr. 1 (2006): 63 - 68, http://dx.doi.org/10.1016/j.cub.2005.10.070
- Kristensen, P. ja Bjerkedal, T. (2007). Sünnikorra ja intelligentsuse seose selgitamine. Science, 316, 1717. https://doi.org/10.1126/ science.1141493.
- Lalumiere, ML, Harris, GT, Quinsey, VL & Rice, ME (1998) Seksuaalne hälve ja vanemate vendade arv seksuaalkurjategijate seas. Seksuaalne väärkohtlemine: ajakiri Research and Treatment 10, 5–15.
- Långström Niklas jt, “Geneetilised ja keskkonnamõjud samasooliste seksuaalsele käitumisele: kaksikute populatsiooniuuring Rootsis”, seksuaalkäitumise arhiiv 39, nr. 1 (2010): 75 - 80, http://dx.doi.org/10.1007/s10508-008- 9386-1.
- Lasco MS jt, „Seksi või seksuaalse sättumuse dimorfismi puudumine inimese eesmises isendis“, Brain Research 936, nr. 1 (2002): 95 - 98, http://dx.doi.org/10.1016/S0006-8993(02)02590-8
- Laumann, EO, Gagnon, JH, Michaels, S. & Michael, RT (1993) AIDSi ja muude haruldaste elanikkonna sündmuste seire: võrgupõhine lähenemine. Tervise ja sotsiaalse käitumise ajakiri 34, 7–22.
- Lauterbach, MD, Raz, S. & Sander, CJ (2001) Vastsündinute hüpoksiline risk enneaegsetel vastsündinutel: sugu ja hingamishäirete raskusaste mõju kognitiivsele taastumisele. Neuropsychology 15, 411-420.
- Lee, JKP jt. (2002) Seksuaalsete kuritegude riskifaktorid. Laste väärkohtlemine Negl. 26, 73 - 92.
- Lee, RM & Silver, RM (2000) Korduv raseduse kaotus: kokkuvõte ja kliinilised soovitused. Reproduktiivmeditsiini seminarid 18, 433–440.
- Lenderking, WR, Wold, C., Mayer, KH, Goldstein, R., Losina, E. & Seage, GR (1997) Laste seksuaalne väärkohtlemine homoseksuaalsete meeste seas. Levimus ja seos ebaturvalise seksiga. Journal of General Internal Medicine 12, 250-253.
- Lenroot RK, Gogtay N, Greenstein DK jt. Aju arengu trajektooride seksuaalne dimorfism lapseeas ja noorukieas. NeuroImage 2007; 36 (4): 1065-1073. doi: 10.1016 / j.neuroimage.2007.03.053.
- LeVay Simon, “Erinevus hüpotalamuse struktuuris heteroseksuaalsete ja homoseksuaalsete meeste vahel”, Science 253, nr. 5023 (1991): 1034 - 1037, http://dx.doi.org/10.1126/science.1887219
- LeVay, S. (2016) .Gay, sirge ja põhjus, miks: Teadus seksuaalsest sättumusest (2nd toim.). Oxford, Suurbritannia: Oxford University Press.
- Lippa Richard A., “Kas 2D: 4D sõrme ja pikkuse suhted on seotud seksuaalse orientatsiooniga? Jah meestele, ei naistele, ”Isiksuse ja sotsiaalpsühholoogia ajakiri 85, ei. 1 (2003): 179 - 188, http://dx.doi.org/10.1037/0022-3514.85.1.179
- Lombardi, CM ja Hurlbert, SH (2009). Ühe sabaga testide vale kirjeldamine ja väärkasutamine. Austral Ecology, 34, 447-468.
- Lykken, D.T., McGue, M., Tellegen, A., "Värbamise kallutatus kaksikuuringutes: kahe kolmandiku kordusreegel" käitumine. Genet. 1987;17:343
- MacCulloch, SI, Gray, NS, Phillips, HK, Taylor, J. & MacCulloch, MJ (2004) Sünnikorraldus seksuaalselt solvavatel ja agressiivselt solvavatel meestel. Seksuaalkäitumise arhiivid 33, 467–474.
- Magnus, P., Berg, K. & Bjerkedel, T. (1985) Pariteedi ja sünnikaalu seos: sensibiliseerimise hüpoteesi testimine. Inimese varajane areng 12, 49–54
- Maguire EA, Gadiani peadirektoraat, Johnsrude IS jt. Navigeerimisega seotud struktuurimuutus taksojuhtide hipokampuses. Ameerika Ühendriikide Riikliku Teaduste Akadeemia toimetised. 2000; 97 (8): 4398-4403.
- Mainardi M, et al. Keskkond, leptiini tundlikkus ja hüpotalamuse plastilisus. Neuraalne plastilisus. 2013. Köide 2013 (2013), artikli ID 438072, 8 lehed http://dx.doi.org/10.1155/2013/438072
- Manikkam, M., Crespi, EJ, Doop, DD, Herkimer, C., Lee, JS, Yu, S., Brown, MB, Foster, DL & Padmanabhan, V. (2004) Loote programmeerimine: sünnieelne testosterooni ületamine viib lammaste loote kasvu aeglustumine ja sünnijärgne järelejõudmine. Endokrinoloogia 145, 790-798.
- Manning JT. (2001) numbrisuhe: osutus viljakusele, käitumisele ja tervisele. Rutgers University Press, London.
- Mantilla K. Bioloogia, mu perse. Seljast väljas: naiste uudiste ajakiri, 5. jaanuar 2004.
- Martin, RM, Smith, GD, Mangtani, P., Frankel, S. & Gunnell, D. (2002) Imetamise ja kasvu vaheline seos: Boydi-Orri kohordi uuring. Lapsepõlve haiguste arhiiv - loote ja vastsündinute väljaanne 87, F193–201.
- Mayer Lawrence S. ja McHugh Paul R., Seksuaalsus ja sugu: avastused bioloogilistest, psühholoogilistest ja ühiskonnateadustest, Uus Atlantis, number 50, sügis 2016, lk. 116. http://www.thenewatlantis.com/sexualityandgender
- Mbugua K. Seksuaalne orientatsioon ja ajustruktuurid: hiljutiste uuringute kriitiline ülevaade. Praegune teaduse köide 84, ei 2 (25 jaanuar 2003), lk. 173-178 (6 lehed). https://www.jstor.org/stable/24108095
- McConaghy, N., Hadzi-Pavlovic, D., Stevens, C., Manicavasagar, V., Buhrich, N. & Vollmer-Conner, U. (2006) Vennalik sünnijärjekord ja heteroseksuaalsete / homoseksuaalsete tunnete suhe naistel ja meestel ... Ajakiri Homoseksuaalsus 51, 161–174.
- McFadden Dennis ja Shubel Erin, “Meeste ja naiste sõrmede ja varvaste suhteline pikkus”, Hormoonid ja käitumine 42, nr. 4 (2002): 492 - 500, http://dx.doi.org/10.1006/hbeh.2002.1833
- Milinski, M. (2006). Suurim histo-ühilduvuse kompleks, seksuaalne valik ja kaaslase valik. Ökoloogia ja süstemaatika aastaülevaade, 37, 159 - 186.
- Mitter C, Jakab A, Brugger PC jt. Inimese loote kommissuraalsete ja sisemiste kapslikiudude in utero traktograafia valideerimine histoloogilise struktuuri tensoanalüüsiga. Neuroanatoomia piirid. 2015; 9: 164. doi: 10.3389 / fnana.2015.00164.
- Morikawa, M., Yamada, H., Kato, EH, Shimada, S., Yamada, T. & Minakami, H. (2004) Korduva raseduse katkemisega naiste normaalse kromosoomikarüotüübiga raseduse katkemises on ülekaalus embrüokadu. Inimese paljundamine 19, 2644-2647.
- Mukherjee, Siddhartha. Geen: intiimne ajalugu. Simon ja Schuster, New York, 2016.
- Mustanski BS, Dupree MG, Nievergelt CM, Bocklandt S, Schork NJ, Hamer DH. Meeste seksuaalse sättumuse genoomne skaneerimine. Hum Genet 2005 Mar; 116 (4): 272-8. Epub 2005 jaan 12.
- New Yorgi põliselanike, 7-10-1995, Gay Gene'i uurimistöö ei allu kontrollile, Chicago Tribune'i John Crewdson paljastab NCI uurija võimaliku teadusliku üleastumise.
- NewsBeat (2015) Vaieldamatu ja teaduslik tõend Michelle Obama on tõesti mees ... NewsBeat Ent. 24.11.2015. newsbeat.co.ke/gossip/irrefutable-scientific-proof-michelle-obama-is-indeed-a-man/
- Newsweek: veebruar 24, 1992 p.49
- NIAAA (2012) alkoholismi perekonna ajalugu. Riiklik alkoholitarbimise ja alkoholismi instituut. https://pubs.niaaa.nih.gov/publications/familyhistory/famhist.htm
- Nimmons D. Seks ja aju. Avastage. 01.03.1994. Discovermagazine.com/1994/mar/sexandthebrain346
- Ngun TC, Guo W, Ghahramani NM, Purkayastha K, Conn D, Sanchez FJ, Bocklandt S, Zhang M, Ramirez CM, Pellegrini M, Vilain E. Seksuaalse orientatsiooni uus ennustav mudel epigeneetiliste markerite abil. Kokkuvõte: Seksuaalse sättumuse uus ennustav mudel epigeneetiliste markerite abil. Esinetud Ameerika inimgeneetika seltsi 2015 aastakoosolekul. Baltimore, Md.
- Nokia MS jt. Füüsiline koormus suurendab täiskasvanud hipokampuse neurogeneesi isastel rottidel, kui see on aeroobne ja püsiv. J Physiol. 2016 (1): 594 (7): 1855-73. doi: 10.1113 / JP271552. Epub 2016 veebr 24.
- Norton R. Kas homoseksuaalsus on päritud? New Yorgi raamatute ülevaade (juuli, 1995). www.pbs.org/wgbh/pages/frontline/shows/assault/genetics/nyreview.html
- Nunez, JL & McCarthy, MM (2003) Soolised erinevused ja hormonaalsed mõjud enneaegse imiku ajukahjustuse mudelis. New Yorgi Teaduste Akadeemia aastakirjad 1008, 281–284.
- Paglia C. Vamps & Tramps: uued esseed. Vintage Books, 1994, lk 71–72
- Parshley Lois. Kas teie geenid võivad teid tappa? Populaarteadus. 28.04.2016. https://www.popsci.com/can-your-genes-make-you-kill
- Paul, JP, et al. (2001) Laste seksuaalse väärkohtlemise mõistmine meeste seksimisega meeste seksuaalse riski võtmise ennustajana: Linna meeste terviseuuring. Laste väärkohtlemine Negl. 25, 557 - 584.
- Paulhus, DL (2008) .Birthorder.InM. Haith (Toim.), Imiku ja varase lapseea arengu entsüklopeedia (köide 1, lk. 204 - 211). San Diego, CA: AcademicPress. https://doi.org/10.13140/2.1.3578.3687.
- Paus T. Aju küpsemise ja kognitiivse arengu kaardistamine noorukieas. Kognitiivsete teaduste suundumused. 2005; 9 (2): 60-68. https://doi.org/10.1016/j.tics.2004.12.008
- Pierik, FH, Burdorf, A., Deddens, JA, Juttmann, RE ja Weber, RFA (2004). Ema ja isa riskifaktorid krüptorhhidiidi ja hüpospaadiate korral: juhtumikontrolli uuring vastsündinud poistel Keskkonna- ja terviseperspektiivid, 112, 1570-1576
- Poasa, KH, Blanchard, R., & Zucker, KJ (2004). Sünnikord transseksuaalidega meestel Polüneesiast: Samoa fa'-afafiini kvantitatiivne uuring. Sugu- ja abieluteraapia ajakiri, 30, 13–23. doi: 10.1080 / 00926230490247110.
- Pulst SM .. Geneetilise ahela analüüs. Arch Neurol. 1999; 56 (6): 667 - 672. doi: 10.1001 / archneur.56.6.667
- Pumberger, W., Pomberger, G. & Geissler, W. (2001) Proktokoliit rinnaga toidetavatel imikutel: panus varases lapsepõlves hematokleesia diferentsiaaldiagnostikasse. Meditsiinijärgne ajakiri 77, 252-254.
- Purcell, DW, Blanchard, R., & Zucker, KJ (2000). Sünnikord tänapäevases geimeeste valimis. Seksuaalkäitumise arhiivid, 29, 349–356.
- Queer valikul. Gayle Madwin http://www.queerbychoice.com/
- Rahman Qazi ja Wilson Glenn D., „Seksuaalne sättumus ja sõrme pikkuse suhe 2nd 4-ni: kas on tõendeid suguhormoonide mõju arengule või arengu ebastabiilsusele?“, „Psychoneuroendocrinology 28, nr. 3 (2003): 288 - 303, http://dx.doi.org/10.1016/S0306-4530(02)00022-7
- Rainer, J. D., Mesnikoff, A., Kolb, L. C., Carr, A., "Homosexuality and Heteroseksuaalsus identsetes kaksikutes" (sh F. J. Kallmanni arutelu) Psychosom Med. 1960;22:251
- Ramagopalan SV, Dyment DA, Handunnetthi L, Rice GP, Ebers GC. Meeste seksuaalse sättumuse uuring genoomis. J Hum Genet. 2010 veebruar; 55 (2): 131-2. http://dx.doi.org/10.1038/jhg.2009.135
- Remafedi G, et al. (1992) Noorukite seksuaalse külgetõmbe demograafia. Lasteravi 89, 714 - 721.
- Rice G jt, “Meeste homoseksuaalsus: seose puudumine mikrosatelliidimarkeritega Xq28”, Science 284, nr. 5414 (1999): 665 - 667, http://dx.doi.org/10.1126/science.284.5414.665
- Richiardi, L., Akre, O., Lambe, M., Granath, F., Montgomery, SM & Ekbom, A. (2004) Sünnikord, siberi suurus ja sugurakkude munandivähi risk. Epidemioloogia 15, 323-329.
- Rind, B. (2001) Gei ja biseksuaalsete noorukite poiste seksuaalkogemused meestega: psühholoogiliste korrelatsioonide empiiriline uurimine mittekliinilises valimis. Seksuaalse käitumise arhiivid 30, 345 - 368.
- Risch N, Squires-Wheeler E, Keats BJ. Meeste seksuaalne sättumus ja geneetilised tõendid. Teadus. 1993, detsember 24; 262 (5142): 2063-5. DOI: 10.1126 / teadus.8266107
- Robinson SJ ja Manning John T., “2ndi suhe 4-nda numbri pikkusesse ja meeste homoseksuaalsusesse”, evolutsioon ja inimese käitumine 21, nr. 5 (2000): 333 - 345, http://dx.doi.org/10.1016/S1090-5138(00)00052-0
- Rohrer, JM, Egloff, B. ja Schmukle, SC (2015). Sünnikorra mõju uurimine isiksusele. Rahvusliku Teaduste Akadeemia toimetised, 112,14224, 14229-10.1073. Https://doi.org/1506451112/pnas.XNUMX.
- Rosario & Scrimshaw 2014, APA käsiraamat, 1. köide, lk 579
- Rosenthal, D., "Geneetiline teooria ja ebanormaalne käitumine" 1970, New York: McGrawHil
- Müük A jt. Keskkond ja aju plastilisus: endogeense farmakoteraapia poole. Psychologica arvustused 2014; Vol. 94, ei 1. https://doi.org/10.1152/physrev.00036.2012
- Salmon, C. (2012). Sünnijärjestus, mõju isiksusele ja käitumisele. V. Ramachandranis (toim) inimese käitumise entsüklopeedia (köide 1, lk 353 - 359). London: Elsevier. https://doi.org/10.1016/B978-0-12-3750 00-6.00064-1.
- Sandberg, DE, Meyer-Bahlburg, HFL, Yager, TJ, Hensle, TW, Levitt, SB, Kogan, SJ & Reda, EF (1995) Hüpospaadiatega sündinud poiste sooline areng. Psühhoneuroendokrinoloogia 20, 693–709
- Sanders AR jt, “Geenivaramu skaneerimine näitab olulist seost meeste seksuaalse sättumusega,” Psühholoogiline meditsiin 45, nr. 07 (2015): 1379 - 1388, http://dx.doi.org/10.1017/S0033291714002451
- Sanders AR jt. Geenivaramu uuring meeste seksuaalse orientatsiooni kohta. Sci rep. 2017; 7: 16950. http://dx.doi.org/10.1038/s41598-017-15736-4
- Satinover J. Homoseksuaalsus ja tõe poliitika. Rakeri raamatud 1996.
- Savic I jt, “Aju vastus oletatavatele feromoonidele homoseksuaalsetes meestes”, Riikliku Teaduste Akadeemia toimetised 102, nr. 20 (2005): 7356 - 7361, http://dx.doi.org/10.1073/pnas.0407998102
- Savin-Williams, R. C & Ream, GL (2006) Puberteetiline algus ja seksuaalne sättumus noorukiea riiklikus tõenäosusvalimis. Seksuaalkäitumise arhiivid 35, 279-286.
- Teadusmeedia keskus (2015). Asjatundjate reaktsioon konverentsiettekandele (avaldamata töö) epigeneetika ja meeste seksuaalse sättumuse teemal. Oktoober 8, 2015. http://www.sciencemediacentre.org/expert-reaction-to-conference-presentation-unpublished-work-on-epigenetics-and-male-sexual-orientation/
- Semenyna, SW, Petterson, LJ, VanderLaan, DP ja Vasey, PL (2017). Samoa androfiilse fa'afafiini ja günifiilse mehe sugulaste reproduktiivse toodangu võrdlus. Seksuaalkäitumise arhiivid, 46, 87–93.
- Serano, J. M. (2010). "Autogünefiilia vastane juhtum." International Journal of Transgenderism. 12 (3): 176–187. doi:10.1080/15532739.2010.514223
- Smith, MJ, Creary, MR, Clarke, A. & Upadhyaya, M. (1998) Sooline suhe ja uniparentaalse disoomia puudumine normaalse karüotüübiga spontaansetes abortides. Kliiniline geneetika 53, 258-261.
- Sorensen, HT, Olsen, ML, Mellemkjaer, L., Lagiou, P., Olsen, JH & Olsen, J. (2005) Meeste rinnavähi emakasisene päritolu; sünnitellimuse uuring Taanis. Euroopa vähi ennetamise ajakiri 14, 185–186.
- Speiser PW jt, “Steroidide 21-hüdroksülaasi puudulikkusest tulenev kaasasündinud neerupealiste hüperplaasia: endokriinseltsi kliinilise praktika juhend,” The Journal of Clinical Endocrinology and Metabolism 95, no. 9 (2009): 4133 - 4160, http://dx.doi.org/10.1210/jc.2009-2631
- Speiser PW, valge arvuti, “Kaasasündinud neerupealiste hüperplaasia”, New England Journal of Medicine 349, nr. 8 (2003): 776 - 788, http://dx.doi.org/10.1056/NEJMra021561
- Stein, Edward, Soovide ebaõnne: seksuaalse orientatsiooni teadus, teooria ja eetika (New York: Oxford University Press, 1999), 145
- Sulloway, FJ (1996). Mässu sündinud: sünnijärjekord, perekonna dünaamika ja loominguline elu. New York: Pantheoni raamatud.
- Swaab DF, “Seksuaalne sättumus ja selle alus aju struktuuris ja talitluses”, Riikliku Teaduste Akadeemia toimetised 105, nr. 30 (2008): 10273 - 10274, http://dx.doi.org/10.1073/pnas.0805542105
- Tannehill B. New Yorker tsiteerib häbiväärselt LGBT-vastast teadlast. Bilerico projekt. 29. juuli 2014. bilerico.lgbtqnation.com/2014/07/new_yorker_shamingly_cites_anti-lgbt_researcher.php
- Taylor, Tim, "Twin Studies of Homoseksuality", bakalaureusetöö, Cambridge'i ülikooli eksperimentaalpsühholoogia osakond, 1992.
- New York Times (2004). Pulmad / pidustused; Dekaan Hamer, Joseph Wilson. Aprill 11, 2004. www.nytimes.com/2004/04/11/style/weddings-celebrations-dean-hamer-joseph-wilson.html (01.12.2017 on kinnitatud)
- Taastumisküla (2017). Miks alkoholism pole pärilik. Taastumisküla. https://www.therecoveryvillage.com/alcohol-abuse/faq/alcoholism-not-hereditary/#gref
- Theodosis DT, et al. Täiskasvanud imetaja hüpotalamuses aktiivsusest sõltuv neuronaalne-gliaalne ja sünaptiline plastilisus. Neuroteaduse maht 57, väljaanne 3, detsember 1993, lehed 501-535. https://doi.org/10.1016/0306-4522(93)90002-W
- Tomeo, ME, Templer, DI, Anderson, S. & Kotler, D. (2001) Heteroseksuaalsete ja homoseksuaalsete isikute lapsepõlve ja noorukiea molestatsiooni võrdlusandmed. Seksuaalkäitumise arhiivid 30, 535-541.
- Tsroadsmap. Kas kategooriliselt vale? A Bailey-Blanchard-Lawrence'i kliiringumajahttp: //www.tsroadmap.com/info/bailey-blanchard-lawrence.html
- Turner, MC, Bessos, H., Fagge, T., Harkness, M., Rentoul, R., Seymour, J. jt. (2005) Ani-HPA-1a põhjustatud vastsündinute alloimmuunse trombotsütopeenia tuvastamiseks võimaliku antennaalse sõeluuringu tulemuste ja kulutasuvuse perspektiivne epidemioloogiline uuring. Vereülekanne 45, 1945 - 1956.
- Rahman Q. http://dx.doi.org/10.1037/0735-7044.117.5.1096
- Van Ombergen, A., Jillings, S., Jeurissen, B., Tomilovskaya, E., Rühl, RM, Rumshiskaya, A., ... Wuyts, FL (2018). Ajukude - muutused kosmoses. New England Journal of Medicine, 379 (17), 1678 - 1680. doi: 10.1056 / nejmc1809011
- VanderLaan, DP, Blanchard, R., Wood, H., Garzon, LC ja Zucker, KJ (2015). Sünnikaal ja kaks võimalikku ema mõju meeste seksuaalsele orientatsioonile: Kliiniline uuring laste ja noorukite kohta suunati soolise identiteedi teenistusele. Arengupsühhobioloogia, 57,25-34. https://doi.org/10.1002/dev.21254.
- Voracek Martin, Manning John T. ja Ponocny Ivo, “Digitaalsuhe (2D: 4D) Austria homoseksuaalsetes ja heteroseksuaalsetes meestes”, seksuaalkäitumise arhiiv 34, nr. 3 (2005): 335 - 340, http://dx.doi.org/10.1007/s10508-005-3122-x
- Wedekind, C., Seebeck, T., Bettens, F., & Paepke, AJ (1995). MHC-st sõltuvad paarilise eelistused inimestel. Proceedings Biological Sciences, 22, 245-249.
- Wellings, K, et al. (1994) seksuaalne käitumine Suurbritannias: seksuaalse hoiaku ja elustiili riiklik uuring. Penguin Books, London
- Valgepea NE. Poistevastane antikeha? Ema immuunsuse hüpoteesi uuesti läbivaatamine. J. biosoc. 2007. doi: 10.1017 / S0021932007001903
- Williams TJ jt, “Sõrme pikkuse suhe ja seksuaalne sättumus”, Nature 404, nr. 6777 (2000): 455 - 456, http://dx.doi.org/10.1038/35006555
- Williams, Zev (20. september 2012). "Raseduse taluvuse esilekutsumine." New England Journal of Medicine. 367:1159–1161. doi:10.1056/NEJMcibr1207279. PMC 3644969
- Wilson JD jt. Seksuaalse arengu hormonaalne kontroll. Teadus 211 (1981): 1278 - 1284, http://dx.doi.org/10.1126/science.7010602
- Witchel Alex. Intervjuu Cynthia Nixoniga. New York Timesi ajakiri. Elu pärast "Seksi". Jaanuar 2012. http://www.nytimes.com/2012/01/22/magazine/cynthia-nixon-wit.html
- Wyndzen, M. H. (2003). Autogynephilia ja Ray Blanchardi valesti suunatud transseksuaalsuse seksitung mudel. Kõik segamini: transsoolise psühholoogiaprofessori vaatenurk elule, soo psühholoogia ja "sooidentiteedi häire". Saadaval: http://www.GenderPsychology.org/autogynpehilia/ray_blanchard/
- Wyre, R. (1990) Miks mehed kuritarvitavad lapsi? Tate, T. (toim.) Lastepornograafia. Methuen, London, lk. 281 - 288.
- Xanthakos, SA, Schwimmer, JB, Aldana, HM, Rothenberg, ME, Witte, DP & Cohen, MB (2005) Allergilise koliidi levimus ja tulemus rektaalsete verejooksudega imikutel: prospektiivne kohordi uuring. Laste gastroenteroloogia ja toitumise ajakiri 41, 16–22.
- Yong E. Ei, teadlased pole gei geeni leidnud. Meedia tõstab esile uuringut, mis ei tee seda, mida ta ütleb. Teadus. Oktoober 10, 2015. https://www.theatlantic.com/science/archive/2015/10/no-scientists-have-not-found-the-gay-gene/410059/
- Zanin E, Ranjeva JP, Confort-Gouny S jt. Inimese normaalse loote aju valgeaine küpsemine. In vivo difusioontensoriga traktograafia uuring. Aju ja käitumine. 2011; 1 (2): 95-108. doi: 10.1002 / brb3.17.
- Zietschi BP. Vennaliku sünnijärjekorra mõju osas tuleb olla ettevaatlik. Arch Sex Behav. 2018. DOI 10.1007 / s10508-017-1086-2
- Zietsch, BP, Verweij, KJH, Heath, AC, Madden, PAF, Martin, NG, Nelson, EC, ... Lynskey, MT (2012). Kas jagatud etioloogilised tegurid soodustavad seksuaalse sättumuse ja depressiooni suhet? Psühholoogiline meditsiin, 42,521 - 532. doi: 10.1017 / s0033291711001577
- Zusman, I., Gurevich, P. & Ben-Hur, H. (2005) Kaks sekretoorset immuunsüsteemi (limaskesta ja barjäär) inimese emakasiseses arengus, normaalne ja patoloogiline (ülevaade). International Journal of Molecular Medicine 16, 127-133.
Isegi identsete kaksikute puhul on homoseksuaalsus kohandatud 1:1. Ja siis on vaja pakkuda vanematele haigestumust, majanduslikke probleeme tervisekvaliteedi säilitamisel ja just nende kontaktide tagamisel, pereprobleeme, kriminaalseid riske ja nii edasi, millega puutub kokku nende laps, kelle õnnest kõik nii väga hoolivad. , kutsudes teda iseseisvalt (?) valima sellist elustiili . Ma üritan seda teha, kuid nad hakkasid mind blokeerima.
Ma arvan, et mõistlik inimene saab aru, et see on ettevõtte huvi. pehmelt öeldes. Inimeste heaolu föderaalse teenistuse spetsialistina ei soovita ma siiralt sellist õnne, mis ei "haise" mitte ainult õnne, vaid ka ülepaisutatud heaolustandardi järgi. Ma ei kujuta ette, et keegi võiks seda tüüpi seksi jaoks välja töötada mingeid hügieenilisi ohutussoovitusi (nali pisaratega...). Muide, ma proovin seda otsida.