
Det meste av materialet nedenfor er publisert i en analytisk rapport. "Retorikken om homoseksuell bevegelse i lys av vitenskapelige fakta". gjør jeg:10.12731/978-5-907208-04-9, ISBN 978-5-907208-04-9
Nøkkelfunn
1. Гипотетический «ген гомосексуализма» не известен, он никем не открыт.
2. Исследования, лежащие в основе заявления о «врожденности гомосексуализма» имеют ряд методологических неточностей и противоречий, и не позволяют сделать однозначных выводов.
3. Даже имеющиеся исследования, цитируемые активистами движения «ЛГБТ+», говорят не о генетической детерминированности гомосексуальных наклонностей, а в лучшем случае о комплексном влиянии, в котором генетический фактор предположительно обусловливает предрасположенность, в сочетании с влиянием окружающей среды, воспитанием и др.
4. Некоторые известные личности среди гомосексуального движения, в том числе учёные, критикуют заявления о биологической предопределенности гомосексуализма и говорят, что он обусловлен сознательным выбором.
5. Авторы методики ЛГБТ-пропаганды «After The Ball» рекомендовали лгать о врождённости гомосексуализма:
«Во-первых, необходимо убедить широкую общественность, что геи являются жертвами обстоятельств, и что они выбирают свою сексуальную ориентацию не более, чем они выбирают свой рост, цвет кожи, таланты или ограничения. Til tross for at tilsynelatende seksuell orientering for folk flest er et produkt av komplekse samspill mellom en medfødt disposisjon og miljøfaktorer i barndommen og tidlig ungdomstid, insisterer vi at for alle praktiske formål bør det vurderes at homofile ble født på den måten.
<..>
Гомосексуалисты ничего не выбирали, никто никогда их не одурачивал и не соблазнял».
Innledning
Argumentet om at homoseksuell tiltrekning er medfødt - den såkalte Hypotesen om den biologiske determinismen om homoseksuell tiltrekning er en av de grunnleggende i "LHBT +" -bevegelsen. Slagordet "Born This Way"1, aktivt spredt i populærkulturen, førte til at mange ikke-spesialister trodde at den biologiske oppfinnelsen av homofili er noe udiskutabel og bevist. Dette stemmer ikke.
De mest pålitelige fakta om homofili indikerer ikke et biologisk, men et sosioøkologisk årsakssammenheng. De siste tiårs innsats for å finne data som ville støtte biologisk teori har bare økt tvilen om at slike data eksisterer.
Oppgaven om den biologiske genesen av homofili er ikke helt spesifikk i seg selv - innenfor dens rammer er det minst to forutsetninger som forklarer mekanismen for den "medfødte naturen" av seksuelle preferanser av samme kjønn: (A) homoseksuell tiltrekning er forårsaket av et "spesielt gen" eller genetisk mutasjon, med andre ord homofili er kodet i humant DNA og overføres fra generasjon til generasjon; (B) homoseksuell tiltrekning er forårsaket av eventuelle avvik under graviditet (hormonelt eller immunforsvar) som angivelig påvirker fosteret i livmoren og resulterer i homoseksuelle preferanser hos babyen.
Dermed vil diskusjonen om den biologiske determinismens hypotese bli delt inn i tre deler. Den første delen vil kritisk undersøke argumentene om forholdet til homofili og gener, den andre delen vil kritisk undersøke argumentene om utviklingen av homoseksuell tiltrekning på grunn av intrauterine hormonelle forstyrrelser. I den tredje delen vil teorien om den autoimmune genesen om homoseksuell tiltrekning bli kritisk undersøkt.

Del en: homofile gener?
Uttalelsen om homofiliens genetiske natur er basert på selektiv presentasjon av noen data og undertrykkelse av andre data blant de aller fleste som ikke har spesialisert kunnskap om genetikk. Vitenskapen kjenner ikke “genet for homofili”, det har aldri blitt identifisert noe sted, selv om det har vært mange forsøk.
Vurder studiene på grunnlag av hvilke LGBT + -aktivister fremmet dette argumentet. For det første er det verdt å kort beskrive med hvilke grunnleggende metoder forskere kan bestemme om en persons eiendom (egenskap) er genetisk bestemt. Disse metodene inkluderer tvillingforskning og molekylærgenetisk analyse.
Tvillingstudier
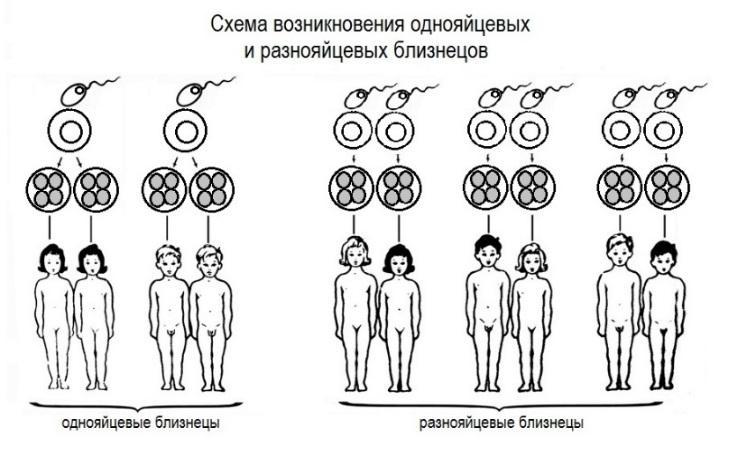
Undersøkelse av identiske tvillinger er en adekvat forskningsmetode for å vurdere om noen egenskaper har genetisk grunnlag. Til å begynne med - hva betyr begrepet “identiske tvillinger”? Slike tvillinger utvikler seg fra det samme befruktede egget, som er delt inn i deler, som separate organismer utvikler seg fra, som er genetiske kopier av hverandre. Genene deres faller sammen på 100%, du kan kalle dem naturlige kloner. Identiske tvillinger kalles også identiske eller monozygote (homozygote) tvillinger. Homoseksuelle tvillinger dannes fra forskjellige egg, befruktet av forskjellige sædceller. Genene deres sammenfaller i gjennomsnitt med 50%, det kan være forskjellig kjønn, høyde, farge på øyne, hår osv. Ikke-identiske tvillinger kalles også ikke-identiske eller dizygotiske (heterozygote) eller dobbelt-tvillinger.
I studien av tvillinger studeres konkordans (tilfeldighet). Konkordans av en egenskap er sannsynligheten for manifestasjonen av en egenskap som begge tvillingene har. Hvis identiteten til noen egenskap hos identiske tvillinger er høy, kan vi konkludere med at denne egenskapen sannsynligvis skyldes genetiske faktorer. Hvis konkordansen til egenskapen til identiske tvillinger ikke overstiger konkordansen i tvillinger til identiske tvillinger, indikerer dette at for dannelsen av denne egenskapen kan det generelle miljøet være en viktigere faktor enn de vanlige genene (Yarygin 2003).
Det er nødvendig å avklare nøyaktig hva samstemthet viser. Det indikerer på ingen måte tilstedeværelsen av noe gen. Konkordansen til en egenskap hos tvillinger indikerer arven til denne egenskapen. Her er det verdt å dvele ved betydningen av ordet “arvelighet” i tvillingstudier. Arv er et mål på hvor mye variabiliteten til en bestemt egenskap i en populasjon (det vil si hvor forskjellig denne egenskapen kan være fra individ til individ) er relatert til variasjonen av gener i en gitt populasjon. I tvillingstudier er imidlertid arvelighet ikke et mål på den genetiske determinismen til en egenskap.
Egenskaper som er nesten fullstendig genetisk bestemt, kan ha veldig lave arvelighetsverdier, mens egenskaper uten tilnærmet genetisk basis kan vise høye arvelighetsverdier. For eksempel er antall fingre - fem på hver lem - hos mennesker nesten helt genetisk bestemt. Men antall fingre hos en person er preget av lav variasjon, og den observerte variabiliteten i de fleste tilfeller forklares av ikke-genetiske faktorer som ulykker, noe som resulterer i en lav arvelighetskoeffisient for egenskapen. Det vil si at hvis du finner tretti par tvillinger der en av dem ikke vil ha fem fingre på hånden, vil det samme antall fingre til den andre broren observeres i et ekstremt lite antall par, om noen.
Derimot kan noen kulturelle trekk være veldig arvelige. Hvis vi for eksempel vurderte å bære øreringer i Amerika i midten av det tjuende århundre, ville vi se at det er preget av en høy grad av arvelighet, siden den var veldig avhengig av kjønn, som igjen er assosiert med tilstedeværelsen av par av XX eller XY-kromosomer, derfor variasjonen i å bære øreringer er sterkt assosiert med genetiske forskjeller, til tross for at dette er mer et kulturelt snarere enn et biologisk fenomen. Hvis du for eksempel ville undersøke tretti par tvillingjenter der en av søstrene har på seg øreringer, så ville i andre tilfeller også andre i øreringer. I dag ville arvelighetskoeffisienten for å bære øreringer være lavere enn i Amerika i midten av det tjuende århundre, ikke fordi det har skjedd endringer i genepuljen til amerikanere, men fordi antallet menn som har øreringer har økt (Blokk xnumx).
En av pionerene innen atferdsgenetikk var en amerikansk psykiater av tysk avstamning, Franz Joseph Kallmann. I en artikkel publisert i 1952 sa han at i 37 par av identiske (monozygote) tvillinger studerte han, hvis en av tvillingene var homoseksuell, så var den andre også homoseksuell, det vil si at grad av konkordans var en svimlende 100% (Kallmann xnumx). Kallmann indikerte ikke nøyaktig hvordan han testet monozygositeten til deltakerne i studien. Forfatteren indikerte heller ikke nøyaktig hvordan han rekrutterte deltakere til studien, mens publikasjonen sa: "søket etter potensielle deltakere var organisert ikke bare ved hjelp av psykiatriske, kriminalomsorgsorganisasjoner og veldedige organisasjoner, men også gjennom direkte kontakter med den underjordiske homoseksuelle verdenen" (Kallmann xnumx). Derfor ble Kallmanns studie hard kritisert (Taylor 1992): Rosenthal indikerte overvekt av personer med psykiatriske problemer blant Kallmann-respondentene (Rosenthal xnumx), Bemerket Likken den uforholdsmessige overvekt av monozygotiske tvillinger i Callamanne-prøven sammenlignet med den generelle befolkningen: (Lykken 1987).

Professor Edward Stein konkluderte med at Kallmann-utvalget “på ingen måte var representativt for den homoseksuelle befolkningen” (Stein xnumx). Dessuten innrømmet Kallmann selv at han anser resultatene for å være noe mer enn en "statistisk artefakt" (Rainer 1960). I statistikk kalles prøver som prøver i Kallmann-studien “praktiske prøver” - de inkluderer valg av objekter i henhold til kriterier som er praktiske for forskeren. Ved å bruke en slik prøve kan man ikke vitenskapelig generalisere, fordi egenskapene til en slik prøve ikke gjenspeiler egenskapene til den generelle populasjonen.
Hvis for eksempel undersøkelsen blir utført i kjøpesenteret tidlig på morgenen bare en dag, representerer ikke resultatene meningene fra andre samfunnsmedlemmer, som det ville være tilfelle hvis undersøkelsen ble utført til forskjellige tider på dagen og flere ganger i uken. Eller hvis du spør kunder i butikken om de vil kjøpe alkohol, så på fredag kveld vil ikke resultatet sammenfalle med resultatene på søndag.
I 1968 undersøkte amerikanske forskere Heston og Shields samstemningen til homofili i identiske 7 tvillingpar. Studiedeltakerne ble funnet i Madsley Twin Register (Heston xnumx). Alle respondentene var psykiatriske pasienter. Forfatterne avslørte samstemthet i identiske tvillinger i 43%. Denne studien ble også kritisert, inkludert av forfatterne selv, på grunn av deltakernes psykiatriske sykdommer og den ekstremt lille utvalgsstørrelsen (Taylor 1992; Heston xnumx).
Studien av Bailey og Pillard
Den neste studien av seksuell tiltrekning blant tvillinger ble utført i 1991 av Michael Bailey fra Northwestern University og Richard Pillard fra Boston University i Amerika (Bailey 1991). De undersøkte samstemningen til homofili hos brødre i ulik grad av slektskap. 56 par identiske tvillinger, 54 par identiske tvillinger, 142 søsken og 57 par stembrødre ble undersøkt2. Tabellen nedenfor viser resultatene fra analysen.
Homoseksuell konkordans
avhengig av graden av forhold (Bailey 1991)
| Forholdstype | Prosentandelen av totale gener | konkordans |
| Identiske tvillinger | 100% | 52% |
| Ikke-identiske tvillinger | 50% | 22% |
| Tvillingbrødre | 50% | 9,2% |
| Stebrødre (ikke pårørende) | Ingen vesentlige likheter | 11% |
Bailey og Pillard uttalte at siden det andre søskenbarnet i et identisk par tvillinger i 52% av tilfellene også hadde homofile preferanser, så "... homofile tendenser skyldes genetisk innflytelse ...".
Bailey og Pillard-studien har, som i tidligere tvillingstudier, grunnleggende problemer. For det første, hvis homofili var genetisk bestemt, ville samstemningen blant identiske tvillinger være 100%, ikke 52%, fordi genene deres er identiske på 100%, og ikke på 52%. I en kommentar til Bailey og Pylards artikkel bemerket Riesch også at nivået av tilfeldigheter blant genetisk fremmede mennesker - halvbrødre - var enda høyere enn blant biologiske ikke-tvillingbrødre, noe som indikerer betydningen av miljøpåvirkning. (Risch 1993). I samsvar med genetikkprinsippene, i tillegg til 100% tilfeldighet av seksuell lyst hos identiske tvillinger, bør prosentandelen av tilfeldigheter hos identiske tvillinger og ikke-tvillinger være høyere enn henholdsvis 22% og 9,2% (se tabell nedenfor).
I tillegg er identiteten til identiske tvillinger (100% av genetisk likhet) forskjellig fra identiteten til identiske tvillinger (50% av genetisk likhet) ved 2.36 ganger, men hvis vi sammenligner identiteten til identiske tvillinger med konkordansen til tvillingbrødrene (50%) forskjellen er: 2.39 ganger, noe som igjen indikerer en mer uttalt innflytelse av miljøet enn genetikk (se tabellen nedenfor).
Sammenligning av konkordans mellom kategorier (Bailey 1991)
| Sammenlign kategorier | Forskjell i genetisk likhet | Forskjellen mellom konkordanser |
| Identiske tvillinger og motsatte tvillinger | Dobbelt så mange vanlige gener |
2.36
|
| Tvillingbrødre og tvillingbrødre | Det er ingen forskjell i prosentandelen av totale gener |
2.39
|
For det andre valgte ikke Bailey og Pillard et vilkårlig utvalg av homoseksuelle. Det vil si at de ikke inkluderte mennesker i studien i henhold til standardene for objektiv akademisk forskning: ikke interessert i resultatene, ikke kjent med hverandre, etc. Som forsker Baron skriver:
“I stedet ble deltakerne rekruttert ved å legge ut annonser i homofile magasiner. Et slikt utvalg av deltakere er veldig tvilsomt, fordi det avhenger av lesertallene til slike magasiner og av motivasjonen til de som gikk med på å delta. Et slikt faktum fører til en forvrengning av resultatene, for eksempel til at antallet homofile tvillinger vil bli overvurdert. Hvorfor? Fordi deltakerne tok hensyn til den seksuelle oppførselen til tvillingbrødrene sine før de ble enige om å delta. Og dette stiller tvil til utvalgets tilfeldighet. For vitenskapelig bevis skal prøven være så tilfeldig som mulig, det vil si at det var nødvendig å inkludere alle tvillingene i undersøkelsen, og deretter foreta en analyse av seksuell atferd ... ”(Baron 1993).
For det tredje, som forskerne Hubbard og Wald skriver i sin analyse:
"... det faktum at konkordansen blant tvillingbrødrene - 22% - mer enn dobler konkordansen blant vanlige brødre - 9,2% - indikerer at grunnen til utviklingen av homofili ikke er genetikk, men miljøet. Den genetiske likheten til heterogene tvillinger ligner faktisk likheten til vanlige brødre. Og hvis miljøfaktorer og oppvekst har så stor innflytelse når det gjelder heterogene tvillinger, er det ikke overraskende at blant identiske tvillinger er påvirkningen av miljøet enda større. Tross alt er den psykologiske oppfatningen av en person som har en identisk tvillingbror uløselig forbundet med denne tvillingen ... ”(Hubbard xnumx).
Forskerne Billings og Beckwiers skrev i sin anmeldelse "... selv om forfatterne tolket funnene som bevis på det genetiske grunnlaget for homofili, tror vi at resultatene tvert imot indikerer at faktorer for oppdragelse og miljø påvirker utviklingen av homofili" (Faktureringer xnumx, s. 60).
Er resultatene fra Bailey og Pillard blitt gjentatt?
Klarte noen å gjenta (gjenskape) resultatene fra Bailey og Pillard - for å finne samstemthet blant identiske tvillinger minst i 52%? I 2000 prøvde Michael Bailey selv å gjenta forskningen sin i en stor gruppe tvillinger i Australia. Konkordansen til homoseksuelle tilbøyeligheter var enda mindre enn i hans første studie. Blant identiske tvillinger var det 20% for menn, og 24% for kvinner, og blant identiske tvillinger - 0% for menn, og 10% for kvinner3 (Bailey 2000).

Kilde: Sally Ryan for The New York Times
I 2010 gjennomførte den svenske epidemiologen Langström en kompleks storstilt studie av seksuell legning hos tvillinger, og analyserte dataene fra flere tusen par identiske og heterofile tvillinger av samme kjønn (Långström 2010). Forskere har identifisert homoseksuelle tendenser når det gjelder eksistensen av seksualpartnere av samme kjønn gjennom livet. De beregnet samstemthet med to parametere: ved tilstedeværelsen av minst en homoseksuell partner i løpet av livet, og av det totale antall homoseksuelle partnere i løpet av livet. Konkordansindikatorene i prøven var lavere enn de som ble oppnådd i begge studiene av Bailey et al. (1991) og (2000). I gruppen av deltakere som hadde minst en partner av samme kjønn, var samstemningen hos menn 18% for identiske og 11% for identiske tvillinger; hos kvinner, henholdsvis 22% og 17%.

Kilde: Karolinska Institut
For det totale antallet seksuelle partnere utgjorde konkordansindikatorene hos menn 5% for identiske og 0% for identiske tvillinger; hos kvinner, henholdsvis 11% og 7%. Hos menn forklares 61% og 66% av varians av miljøfaktorer som bare påvirker en tvilling av et par, mens varians ikke i det hele tatt er forklart av miljøfaktorer som er vanlige for tvillinger. Unike miljøfaktorer utgjorde henholdsvis 64% og 66% spredning, mens generelle miljøfaktorer utgjorde henholdsvis 17% og 16% (Långström 2010).
I 2002 gjennomførte forskerne Peter Birmen fra Columbia University og Hannah Bruckner ved Yale University of America en omfattende og representativ studie med et stort antall deltakere (Bearman 2002).

Kilde: hannahbrueckner.com
De fikk enda mer ubetydelige nivåer av samstemthet av homofile tilbøyeligheter: 6,7% i par av identiske tvillinger, 7,2% i forskjellige identiske tvillinger og 5,5% i vanlige brødre. Birmen og Bruckner konkluderte med at de ble funnet:
"... betydelige bevis til fordel for modellen for sosialisering på individnivå ..., våre resultater antyder at å oppdra barn på prinsippet om kjønnsnøytralitet, uten å tydelig etablere kjønnet til barnet, har innvirkning på dannelsen av homoseksuelle tilbøyeligheter ..." (Bearman 2002).
I motsetning til verkene som nettopp ble gjennomgått, gjennomførte psykiateren Kenneth Kendler og hans kolleger en stor tvillingstudie ved bruk av en sannsynlig prøve bestående av 794 tvillingspar og 1380 vanlige brødre og søstre (Kendler xnumx). Forfatterne konkluderte med at funnene deres "antyder at genetiske faktorer kan ha en sterk effekt på seksuell legning." Studien var imidlertid ikke tilstrekkelig til å trekke så alvorlige konklusjoner om graden av påvirkning av gener på seksualitet: I alt, i 19 av 324 par identiske tvillinger, ble en person med homoseksuelle tilbøyeligheter identifisert, mens i 6 av 19-par var homoseksuelle tilbøyeligheter konkordante (observert i andre bror); minst én person med homoseksuelle tendenser ble funnet hos 15 av 240-par av tvillinger av samme kjønn, mens 2 av 15-par var samstemte. Det faktum at bare i 8 av 564 tvillingpar sammenføyte homoseksuelle tilbøyeligheter (1,4%) muligheten for å bruke disse resultatene for en seriøs sammenligning av identiske og ikke-identiske tvillinger.
Det må huskes at identiske tvillinger er omgitt av nesten det samme miljøet - tidlig kjærlighet, forhold til andre barn, etc. - i sammenligning med ikke-identiske tvillinger og vanlige brødre og søstre. Siden identiske tvillinger ligner utseende og karakter, er den samme holdningen til dem mer sannsynlig enn identiske tvillinger og vanlige brødre og søstre. I noen tilfeller kan derfor en høyere konkordansekoeffisient forklares med miljømessige og ikke genetiske faktorer.

Professor Kenneth Kendler.
Kilde: Virginia Commonwealth University.
I følge psykiater Jeffrey Satinover (Satinover xnumx) faktorer som omfattende påvirker dannelsen av en seksuell atferdstype til en person kan deles inn i fem kategorier:
1) intrauterine (prenatal) effekter, for eksempel konsentrasjonen av hormoner;
2) ekstrauterine (postnatal) fysiske effekter som traumer og virusinfeksjoner;
3) ekstrauterine opplevelser, som familieinteraksjoner, utdanning;
4) ekstrauterin opplevelse, for eksempel den forsterkende effekten av stereotyp repetitiv atferd;
5) valg.

Kilde: ihrc.ch
Fraværet av 100% samstemthet i identiske identiske tvillinger antyder ikke bare at påvirkningen av genetiske faktorer er ubetydelig, men også at ikke-genetiske faktorer ikke utelukkende kan være intrauterin. Tross alt, hvis dette var slik, ville konkordans fortsatt være nær 100%, siden identiske tvillinger påvirkes av de samme faktorene i det intrauterine miljøet ”(Satinover xnumx, s. 97).
Hvis gener spiller en rolle i å forme folks disposisjon for visse seksuelle ønsker og atferd, lar alle disse studiene oss med tillit si at dette emnet ikke blir utmattet av påvirkning fra genetiske faktorer. Oppsummere tvillingenes forskning kan vi trygt konkludere med at vitenskapen ikke har bevist at seksuell lyst generelt og homofile tilbøyeligheter spesielt bestemmes av menneskelige gener.
Molekylærgenetiske studier
Å studere spørsmålet om genetikkens deltakelse i dannelsen av homoseksuelle tilbøyeligheter og om mulig graden av denne deltakelsen, har vi så langt undersøkt studier der den genetiske arven til en egenskap (i det spesielle tilfelle av homoseksuell tiltrekning) bestemmes av klassisk genetikk, men de satte ikke oppgaven med å bestemme hvilken spesifikke gener er ansvarlige for denne egenskapen. Samtidig kan genetikk studeres ved hjelp av den såkalte. molekylære metoder som gjør det mulig å bestemme hvilke spesifikke genetiske varianter som er assosiert med fysiske eller atferdsegenskaper.
Dean Haymer Study
Et av de første forsøkene på å utføre en molekylær genetisk analyse av homoseksuelle tilbøyeligheter ble gjort av Dean Haymer og hans kolleger ved National Institute of Health i Maryland, i Amerika (Hamer 1993). Haymer undersøkte familier med identiske mannlige tvillinger, der minst en av tvillingene hadde attraksjon av samme kjønn. Blant det totale antall familier identifiserte Haymer 40, der den homoseksuelle broren hadde en annen bror som også var homoseksuell, og undersøkte deres DNA for lignende nettsteder. En lignende studie kalles “linked inheritance research” - på engelsk “genetisk linkage study”.
I studien av koblet arv gjøres følgende: i en gruppe av personer som har et kjent kjent attributt, blir det utført en analyse for tilstedeværelse av lignende DNA-seksjoner - de kalles markører. Hvis det viser seg at i gruppen av individer det høye antallet markører er lokalisert i samme DNA-region, kan det antas at alle disse markørene er arvet “sammen” - koblet - det vil si at de kan være en del av noe gen (Pulst 1999).
Haymer sa at i 33-par fra 40 har homoseksuelle brødre det samme kjønnsområdet på X-kromosomet, som han kalte "Xq28." Heimer konkluderte med at Xq28-regionen inneholder gener for homoseksuelle tendenser.

forfattere av kontroversielle artikler -
på en konferanse om genetikk og seksualitet,
Mai 1995 (Finn 1996)
Først av alt skal det bemerkes at Haymers resultater ofte blir fortolket feil. Mange mennesker tror at Haymer fant en identisk DNA-region - Xq28 - i alle 33-par, i alle 66-menn, men faktisk viste det seg at nukleotidsekvensene til Xq28-regionen var identiske mellom brødre i hvert tvillingpar, og Xq28-sekvensen i alle par var ikke identisk - Haymer fant ikke det beryktede "homofile genet."
Denne studien har en rekke betydelige ulemper. Haymer sjekket ikke sammenfallet av Xq28 hos tvillingpar med heterofil tiltrekning, men bare blant homoseksuelle (Byne xnumx). Hvis han ikke fant dette nettstedet blant heterofile brødre, men bare blant homoseksuelle, ville dette snakke om et resultat til fordel for hans konklusjon. Hvis han imidlertid hadde oppdaget Xq28 blant sine heterofile brødre, ville konklusjonene hans ha fått null verdi (Horton xnumx). Som forskere Fausto-Sterling og Balaban bemerket, inneholder Heimer-prøven også en ufullstendig mengde data: av 40 tilfeller, bare i 15 DNA-heterozygositetskarakteristikker ble direkte målt; i de resterende 25 tilfellene ble dataene indirekte beregnet (Fausto-Sterling 1993). Bare i 38% av tilfellene målte Heimer et al direkte nivået av heterozygositet til mors mors kromosom, og i 62% beregnet de bare det basert på tilgjengelige databaser.
Det skal nevnes følgende episode relatert til publikasjonen av årets Haymer 1993. I 1995 publiserte magasinet New York Native en artikkel med tittelen "Forskning på" genene "for homofili stod ikke på prøve: journalisten John Krudson fra Chicago Tribune avdekket en sannsynlig vitenskapelig forfalskning begått av en forsker" (Chicago Tribune 1995). Artikkelen indikerer at Haymers arbeid ble kritisert hardt av forskjellige lærde for at Haymer ikke foretok en verifikasjonskontroll for tilstedeværelsen av Xq28 blant heterofile brødre. Kritikere inkluderte anerkjente biologer og genetikere Richard Levontin og Ruth Hubbard fra Harvard University (Chicago Tribune 1995). I samme artikkel heter det videre at Federal Bureau of Ethics fra National Institute of Health studerer klagen til en av de unge ansatte på Heimer-laboratoriet, hvis navn ikke er kjent, som rapporterte manipulasjonen av resultatene gjort av Heimer i sin studie: ifølge uttalelsen fra denne offiseren, Heimer bevisst ekskluderte resultatene som indikerer usaklighet i teorien om genetisk forhåndsbestemmelse av homoseksuelle tilbøyeligheter fra publikasjonenChicago Tribune 1995). Noen måneder etter publiseringen av artikkelen i New York Native, publiserte bladet Scientific American en annen artikkel som bekreftet faktum og grunn for etterforskningen av Federal Ethics Bureau mot Heimer (Horgan xnumx, s. 26). Nasjonalt helseinstitutt avslørte ikke resultatene av etterforskningen, men Haymer ble senere overført til en annen avdeling. Det skal også bemerkes at Haymer utførte sin forskning på “homoseksualitetsgenet” ved å bruke en bevilgning, som faktisk ble tildelt for å studere Kaposis sarkom, en hudkreft som ofte rammer homoseksuelle pasienter med aids (Mukherjee xnumx, s. 375). Gyldigheten av Haymers publikasjon var avhengig av om et uavhengig forskerteam kunne oppnå de samme resultatene. Dette skjedde ikke.

Replikbarhet av Haymer-resultater
I 1999 utførte en gruppe forskere fra University of Western Ontario, ledet av en forsker ved navn Rice, en lignende (ved bruk av "genetisk kobling" -metode) -studie blant 52 homofile menn (Ris xnumx). Forfatterne klarte ikke å gjenta resultatene oppnådd av Haymer og konkluderte: "resultatene av vår studie avslørte ikke noe bevis for en sammenheng mellom mannlig homofili og gener."
Deretter ble det i 2005 utført en ny studie med Dean Haymer (Mustanski Xnumx). Forfatterne fant ikke et statistisk signifikant forhold mellom Xq28 og tilbøyeligheter til homofile, men uttalte at de fant en "interessant korrelasjon" for andre nettsteder (på kromosomene 7, 8 og 10).
Disse resultatene kunne imidlertid ikke gjentas i en annen studie i 2009, da en gruppe forskere fra Oxford i England og University of Ontario i Canada gjennomførte en studie av 55-familier der det var homofile menn: genetisk materiale ble samlet inn fra 112-deltakere og et genomomfattende søk etter assosiasjoner ble utført med inkludering av 6000-genmarkører (Ramagopalan 2010). Analysen avdekket ikke et statistisk signifikant forhold mellom genetiske markører og homofili.
I 2015 uttalte en gruppe forfattere fra forskjellige vitenskapelige sentre i Amerika, ifølge et genomomfattende søk etter assosiasjoner, at de fant et betydelig forhold for nettstedet på 8-kromosomet og mindre signifikant for Xq28 (Slipemaskiner xnumx). I konklusjonene i artikkelen deres innrømmet forfatterne at "den genetiske effekten på homoseksuelle tendenser langt fra er avgjørende ... mest sannsynlig er denne effekten del av en multifaktoriell årsak."
I 2017 brukte den samme gruppen av forfattere en mer moderne og nøyaktig metode som kalles genomomfattende søk etter assosiasjoner4. Letingen etter genomomfattende assosiasjoner er basert på bruk av genomsekvenseringsteknologi (lesing av informasjon fra DNA) for å bestemme de spesifikke funksjonene til DNA som kan være assosiert med egenskapen som undersøkes. Forskere undersøker millioner av genetiske varianter i et stort antall individer med et felles attributt, og individer som ikke har dette attributtet, og sammenligner hyppigheten av genetiske varianter mellom begge grupper. Det antas at de genetiske variantene som er mer vanlig blant eiere av en egenskap enn blant de uten det på en eller annen måte er relatert til denne egenskapen. Denne gangen ble det funnet statistisk signifikante sammenhenger for regioner på kromosomene 13 og 14 (Slipemaskiner xnumx).

En studie av Sanders og kolleger (2017) fant ikke et gen for homoseksuelle tendenser, og beviste ikke deres genetiske tilstand (forfatterne selv unngår det), og bekreftet heller ikke resultatene fra årets Haymer 1993, som la grunnlaget for en lang krangel med homoseksualitetsgener. En av konklusjonene i denne publikasjonen var antagelsen om at alle de ovennevnte genetiske variantene kan påvirke predisposisjon tilbøyeligheter til homofile (Slipemaskiner xnumx, s. 3).
Francis Collins, prosjektleder for avkoding av det menneskelige genom, skriver følgende:
"Sannsynligheten nær 20% for at en identisk tvilling av en homoseksuell mann også vil være homoseksuell (sammenlignet med 2 - 4% i den generelle befolkningen) indikerer at seksuell orientering påvirkes av gener, men ikke innlemmet i DNA, og eventuelle gener involvert. representere en predisposisjon, men ikke en forhåndsinngått konklusjon ... ”(Collins 2006).
En spesielt stor studie om metoden for genomomfattende søk etter assosiasjoner, som hadde som mål å bestemme de genetiske variantene forbundet med homoseksuelle tilbøyeligheter, ble presentert på den årlige konferansen til American Society of Human Genetics i 2012 (Drabant 2012). Som et resultat av et genomomfattende søk ble det ikke funnet signifikante forhold for homofile tilbøyeligheter hos begge kjønn. Samtidig ble mange tusen individer fra firmadatabasen 23andMe undersøkt.
Forfattere av de nyeste og største forskning på genetikken til homofili fortalte om resultatene:
"Det er praktisk talt umulig å forutsi en persons seksuelle atferd basert på deres genom,"
sier Ben Neal, professor i avdelingen for analytisk og translasjonell genetikk ved Massachusetts General Hospital, som jobbet med studien.
Ifølge professor David Curtis ved University of California Genetics Institute,
"Det er ingen kombinasjon av gener i den menneskelige befolkningen som har en betydelig innvirkning på seksuell legning. Det er praktisk talt umulig å forutsi en persons seksuelle oppførsel basert på deres genom."
epigenetikk
I 2015, en gruppe forskere fra University of California, presenterte Los Angeles et sammendrag på en konferanse fra American Society of Human Genetics5som hevdet at forskere var i stand til å identifisere seksuelle preferanser basert på epigenetiske markører med en nøyaktighet på 67% (Ngun et al. 2015). For å tiltrekke maksimal oppmerksomhet til arbeidet sitt, organiserte forfatterne til og med en pressemelding som involverte pressen (ASHG 2015). Nyhetene spredte seg umiddelbart over overskriftene til mainstreamavisene, til tross for studiens åpne motstridende natur og den tvilsomme formidlingsmetoden (Yong xnumx).
Epigenetikk er en vitenskap som studerer fenomener der uttrykket av gener endres på grunn av påvirkning fra mekanismer som ikke påvirker endringen i DNA-sekvensen i gener. Epigenetiske prosesser er med andre ord de prosessene der andre faktorer påvirker graden av genuttrykk (det vil si kroppens fysiologiske egenskaper). Den romlige konfigurasjonen av et DNA-molekyl kan påvirke genuttrykk (ekspresjon), og denne konfigurasjonen bestemmes av spesielle regulatoriske proteiner, enzymer assosiert med DNA. En av mekanismene for påvirkning er DNA-metylering. Kombinasjonen av regulatoriske proteiner og DNA kalles en epigenetisk markør.
Unge og kolleger uttalte at hovedhensikten med studien var å teste muligheten for å bestemme den seksuelle orienteringen til et individ av epigenetiske markører. For dette formål studerte de DNA-prøver av 37 par identiske tvillingbrødre, i hvert av disse parene var en bror homoseksuell, og 10 par identiske tvillingbrødre, der begge brødrene var homoseksuelle. Som det fremgår av sammendraget, studerte forskerne en rekke klassifiseringsmodeller (heterofile kontra homoseksuelle) ved å bruke FuzzyForest datamaskinstatistikkalgoritme og valgte til slutt den best-presterende modellen inkludert 5 epigenetiske markører som riktig klassifiserte objekter i 67% av tilfellene. Forfatterne antydet at seksuelle preferanser kontrolleres av 5 epigenetiske markører. Imidlertid forårsaket en slik tolkning mildt sagt en mengde kritikk fra eksperter (Science Media Center 2015, Virkelig xnumx, Yong xnumx, Gelman 2015, Briggs 2015). Metodikken (ekstrem lav prøvekraft, tvilsom statistisk tilnærming med høy risiko for falske positive resultater osv.) Og dens tolkning forårsaket stor tvil. John Grillie fra Center for Epigenomics ved Albert Einstein College of Medicine bemerket, og kommenterte hypen rundt studien av Ngun og kolleger:
“… Uten å snakke personlig om ham eller kollegene, men hvis vi ønsker å bevare dette vitenskapsområdet, kan vi ikke lenger tillate dårlig epigenetisk forskning. Med "dårlig" mener jeg ufortolket. ... "(Virkelig xnumx).

Til slutt ble objektiviteten til anmelderne som hoppet over denne CVen for presentasjon på konferansen til og med stilt spørsmål ved, og artikkelen ble selvfølgelig aldri publisert noe sted.
Hvorfor er resultatene fra molekylærgenetiske studier så selvmotsigende - variabel og variabel?
Genetikkens begrensede rolle
Bevis for genetisk karakter av homofile tilbøyeligheter er uholdbart. Vitenskapen kjenner ikke "homoseksualitetsgenet." På begynnelsen av dette århundre ble et stort internasjonalt prosjekt "Human Genome Project" lansert - Human Genome Project. Innenfor dens rammer ble kompilering av humane genetiske kart utført - hvilket gen, på hvilket kromosom ligger, hvilke proteiner det koder osv. Hvem som helst kan sjekke - ingen homoseksualitetsgener er indikert der (Menneskelige genomressurser hos NCBI).
Dette er hva Mayer og McHugh skriver i sitt arbeid:
“Som gjentatte ganger blitt bekreftet i forhold til atferdsegenskapene til en person, er en genetisk faktors innflytelse på tendensen til homoseksuelle tilbøyeligheter eller atferdsmønstre mulig. Den fenotypiske manifestasjonen av gener avhenger vanligvis av miljøfaktorer - et annet miljø fører til dannelse av forskjellige fenotyper selv for de samme genene. Derfor, selv om noen genetiske faktorer påvirker homoseksuelle tendenser, påvirkes også seksuelle preferanser og tendenser av en rekke miljøfaktorer, inkludert sosiale belastningsfaktorer som psykologisk og fysisk vold og seksuell trakassering. For å få et mer fullstendig bilde av dannelsen av seksuelle interesser, ønsker og stasjoner, er det nødvendig å ta hensyn til faktorene utvikling, miljø, erfaring, samfunn og vilje. (For eksempel har sosiale genetikere registrert en indirekte rolle av gener i atferd med jevnaldrende, noe som indikerer at en persons utseende kan påvirke aksept eller avvisning i en bestemt sosial gruppe (Ebstein 2010).
Moderne genetikk vet at gener påvirker rekkevidden til et individ og hans motivasjon og følgelig indirekte påvirker atferd. Selv om gener dermed kan lure en person til en viss oppførsel, er deres evne til direkte å kontrollere handlinger, uavhengig av et bredt spekter av andre faktorer, veldig, veldig usannsynlig. Deres innflytelse på atferd er mer subtil og avhenger av virkningen av miljøfaktorer ... "(Mayer 2016).
Medfødte faktorer som kan påvirke orientering inkluderer kvaliteter av temperament som en mild og sårbar karakter, økt emosjonell følsomhet, sjenanse, passivitet, etc. Forskere selv, hvis resultater brukes i retorikken til LHBT + aktivister - bevegelser, tør ikke påstå at homofili bestemmes av gener, i beste fall tror de at attraktivitet av samme kjønn er assosiert med en kombinasjon av biologiske og miljømessige faktorer, der sistnevnte spiller en viktig rolle . At homofili er "medfødt", hører vi hovedsakelig i Hollywood-filmer, rangering av talkshow, sanger eller i kommentarer på sosiale nettverk. Men i det vitenskapelige samfunnet er det faktisk ikke en eneste samvittighetsfull forsker som vil si at han har funnet en genetisk eller noen annen biologisk årsak til homoseksuell tiltrekning.
Studier som hadde som mål å prøve å finne ut om gener (spesielt på Xq28-nettstedet) eksisterer assosiert med seksuell lyst av samme kjønn. Utarbeidet av V. Lysov (2018)
| Kilde og sampling |
metode analyse |
Resultater i henhold til publisering | Er det bevis på et forhold mellom Xq28-markører og homofili? | Andre resultater |
| Dean Hamer et al. 1993 40-familier, som hver bestod av en homofil proband og homoseksuelle valgt blant hans slektninger |
koblede arvstudier | i 33 tilfeller fra 40-familier falt de genetiske markørene på stedet q28 av kromosom X sammen | betingetImidlertid kritiseres metoder og tolkning av kolleger: Baron 1993; pool 1993; Fausto-Sterling et al. 1993; Skarp 1993; Byne xnumx; McLeod 1994; Norton 1995, Haymer ble selv mistenkt for forfalskning: Horgan xnumx | - |
| Jennifer Macke et al. 1993 36 familier, som hver bestod av en homofil proband og hans slektninger, blant dem var minst en homoseksuell bror |
søk etter kandidatgener - androgenreseptorgen (X-kromosom) | ingen statistisk signifikante sammenhenger ble funnet i utvalget | - | ingen forbindelse med androgenreseptorgenet (X-kromosom) |
| Stella Hu et al. 1995 (vitenskapelig gruppe Dean Hamer) 33 familier, som hver bestod av en homofil proband og hans slektninger, blant dem var minst en homoseksuell bror |
koblede arvstudier | i 22 tilfeller fra 32-familier falt de genetiske markørene på stedet q28 av kromosom X sammen | betingetse Hamer 1993 | - |
| George Rice et al. 1999 46 familier, som hver bestod av en homofil proband og hans slektninger, blant dem var minst en homoseksuell bror |
koblede arvstudier | genetiske markører lokalisert på q28-regionen til kromosom X stemte ikke overens | ikke | - |
| Michael DuPree et al. 2004 (vitenskapelig gruppe Dean Hamer) 144 familier, som hver bestod av en homoseksuell proband som hadde minst en homoseksuell bror |
søk etter kandidatgener - aromatase gen CYP15 (15 kromosom) | ingen statistisk signifikante sammenhenger ble funnet i utvalget | - | ingen forbindelse med aromatasegenet CYP15 (15-I kromosom) |
| Mustanski et al. 2005 (vitenskapelig gruppe Dean Hamer) 146-familier (inkludert familier fra studiene av Hamer 1993 og Hu 1995), som hver bestod av et homoseksuelt proband som hadde minst en homoseksuell bror |
genomomfattende studie av koblet arv | en statistisk signifikant assosiasjon med en markør på 7-kromosomet ble funnet i prøven, og ifølge forfatterne, "nærhet til kriterier av sannsynlig betydning" for markører på 8 og 10-kromosomene. | ikke | kommunikasjon med markører på 7-kromosomet i henhold til kriteriene fra Lander og Kruglyak (1995), den beste indikatoren for LOD* tilsvarte xnumx |
| Sreeram Ramagopalan et al. 2010 (George Rice Science Team) 55 familier, som hver bestod av en homoseksuell proband som hadde minst en homoseksuell bror |
genomomfattende studie av koblet arv | ingen statistisk signifikante sammenhenger ble funnet i utvalget | ikke | ingen forbindelser ble funnet med markører på 7-kromosomet i henhold til kriteriene fra Lander og Kruglyak (1995) |
| Binbin Wang et al. 2012 en gruppe av Xnumx homoseksuelle menn og en kontrollgruppe av Xnumx heterofile menn |
søk etter kandidatgener - sonisk pinnsvin (SHH) -gen (7-kromosom) | ingen statistisk signifikante sammenhenger ble funnet i utvalget | - | fant en statistisk signifikant intergruppeforskjell i forholdet mellom mutasjoner i posisjonen rs9333613 av genet, som ble tolket av forfatterne som "tilstedeværelsen av en mulig sammenheng mellom mutasjoner i genet og attraksjon av samme kjønn" |
| Emily Drabant et al. 2012 7887 menn og 5570 kvinner (ikke relatert til slektskap) som har blitt identifisert som å ha sexlyst og selvidentifisering ifølge Klein |
full genomforeningssøk | ingen statistisk signifikante (5 × 10 - 8) assosiasjoner ble funnet i utvalget | ikke | ingen statistisk signifikante assosiasjoner ble funnet |
| Sanders et al. 2015 384 familier, som hver bestod av en homoseksuell proband som hadde minst en homoseksuell bror |
genomomfattende studie av koblet arv | en statistisk signifikant assosiasjon med en markør på 8-kromosomet og en sannsynlig assosiasjon med Xq28 ble funnet i prøven | betinget: i henhold til kriteriene fra Lander og Kruglyak (1995), var de beste LOD-indikatorene for Xq28-markører lik 2,99, som tilsvarer den antatte verdien ("antydende betydning") | kommunikasjon med markører på 8-kromosomet i henhold til kriteriene fra Lander og Kruglyak (1995); den beste LOD-poengsummen var 4,08 |
| Sanders et al. 2017 en gruppe av 1077 homoseksuelle menn og 1231 heterofile menn (de samme forsøkspersonene som Sanders et al. 2015) |
full genomforeningssøk | ingen statistisk signifikante (5 × 10 - 8) assosiasjoner ble funnet i utvalget | ikke | ingen statistisk signifikante sammenhenger ble funnet. Forfatterne bemerket at verdier som nærmet seg signifikant ble oppnådd for markører på kromosomene 13 og 14 |
* LOD = multipoint logaritme av oddsen se Nyholt DR. Alle LOD-er er ikke opprettet like. Am J Hum Genet. 2000 Aug; 67 (2): 282 - 288. http://doi.org/10.1086/303029. Den statistisk signifikante LOD i genetisk forskning er ≥3,
Som en av de amerikanske bloggerne treffende uttrykte det, "... forsøk på å biologisk forklare homofili er lik iPhones - en ny vises hvert år ..." (Allen 2014). Til slutt, sannsynligvis, fra synspunktet til promotørene av homoseksuelle tilbøyeligheter, slagordet "Sannsynligvis født disponert"6 har en helt annen propagandaeffekt.

Det er gjort forsøk på å oppdage “alkoholismegenet” (Restitusjonsbyen 2017; NIAAA 2012), og "morderen genet" (Davis 2016; Persille xnumx) derimot, som i tilfellet med "homoseksualitetsgenet", ble det ikke funnet noen bevis til fordel for påstanden om at "slike er født". En tilstrekkelig person ville ikke ha ideen på den ene siden til å rettferdiggjøre alkoholisme og drap med påvirkning av gener - tross alt, disse fenomenene bestemmes av valg, ikke forhåndsbestemt. Pioneren til historien med "homoseksualitetsgenet" Dean Haymer har åpenbart et utmerket kommersielt talent, og dyktig opptrer innenfor rammen av offentlig mote. Etter å ha ventet kort tid etter publiseringen av årets 1993-artikkel, publiserer Haymer boken "The Science of Passion: the Search for Homosexuality Genes and Behavioural Biology", som gjorde en plask blant LGBT + -bevegelsenHamer 1994) og brakte ham betydelig fortjeneste. Ti år senere gir Haymer en ny sensasjon ved å gi ut en bok med tittelen “Guds gen: Hvordan tro blir forhåndsbestemt av våre gener” (Hamer 2004), der han uttrykte sin mening om at troende nærmest er genetiske mutanter (V.L .: det er morsomt å observere en slik selektivitet med hensyn til to genetiske hypoteser: den påståtte genetiske tilstanden til homofile tilbøyeligheter blir presentert i et positivt lys, som en gitt , og den påståtte forbindelsen mellom gener og religion er negativ, som en mutasjon.) Naturligvis er det ikke funnet noen bekreftelse på Heimers hypoteser i dag, men teorien hans ble også veldig varmt mottatt i LGBT + -samfunnet. Det amerikanske magasinet Time publiserte til og med et spesielt omslag for denne anledningen.

Deretter forlot Dean Haymer vitenskapen og fokuserte på sosiopolitiske aktiviteter: sammen med sin "mann" Joseph Wilson (The New York Times 2004) han grunnla filmstudioet "QWaves", og spesialiserte seg på produkter med fokus på bevegelsen "LGBT +" (Huffpost 2017).
Den berømte biologen og popularisereren av vitenskapen Richard Dawkins karakteriserer filosofisk hypotesen om genetisk determinisme om homofili:
“… Noen ting som er betinget av miljøet er enkle å endre. Andre er vanskelige. Tenk på hvor dypt vi er koblet til aksenten i barndommen vår: en voksen innvandrer blir stemplet som en utlending hele livet. Det er en mye strengere determinisme her enn i virkningen av de fleste gener. Det ville være interessant å vite den statistiske sannsynligheten for at et barn som har blitt utsatt for en viss påvirkning av miljøet, for eksempel religiøs utdannelse i et kloster, senere vil kunne kvitte seg med denne innflytelsen. Det ville være like interessant å vite den statistiske sannsynligheten for at en mann med et bestemt gen på X-kromosomet i Xq28-regionen vil være homofil. Den enkle demonstrasjonen av at det er et gen som "fører" til homofili, etterlater spørsmålet om betydningen av denne sannsynligheten nesten helt åpen. Gener har ikke monopol på determinisme ... "(Dawkins xnumx, s. 104).
En av de mest fremtredende figurene i russisk sexologi, professor Georgy Stepanovich Vasilchenko, som snakker om årsakene til dannelsen av homoseksuelle tilbøyeligheter, peker på følgende:
“… Forstyrrelser i hjernedifferensiering og hormonforskyvninger bestemmer imidlertid ikke dannelsen av homofil tiltrekning, men blir grunnlaget for forvrengninger av seksuell identitet og kjønnsrolleatferd, noe som øker risikoen for homofili. Nevroendokrin støtte er bare en energisk komponent av libido. Dannelsen av homofili er også tilrettelagt av etiologiske faktorer og patogenetiske mekanismer som ligger i perversjoner generelt ... "(Vasilchenko 1990, s. 430).
Hypotesen om den genetiske faktoren til mannlig homoseksualitet, som gir en evolusjonær fordel for kvinner
Det er verdt å nevne den bisarre hypotesen til italienske forskere, som ifølge dem "Passer ikke til noen eksisterende genetisk modell for homofili". Antakelsen om at homoseksualitet er forårsaket av gener er i strid med prinsippet om naturlig seleksjon, hvorved antallet genbærere som hindrer implementeringen av de heterofile funksjonene som er nødvendige for produksjon av avkom, bør reduseres jevnlig til det forsvinner helt. Imidlertid, som vist statistikk, øker antall mennesker som anser seg som homoseksuelle for hver generasjon. Årsaken er klar: homoseksualitet er ikke genetisk drevet, men å ikke ville stille opp med den åpenbare Camperio-Ciani og kolleger kom med en sofistikert forklaring som skulle takle "Darwin-paradokset." Deres hypotese antyder eksistensen av en viss "X-kromosomal faktor", som, overført gjennom mors linje, kan øke androfili (seksuell tiltrekning til menn) hos begge kjønn, og dermed føre til økt fruktbarhet hos kvinner, og kompensere for den reduserte fruktbarheten hos menn (Camperio-Ciani 2004).
Denne hypotesen kan hevde en viss grad av troverdighet hvis forskere fant passende kompensasjonsnivåer - for eksempel hvis en mor med heterofile avkom hadde et 2-barn, og en mor med et homoseksuelt avkom hadde 4. Faktisk viste forskjellen seg å være ubetydelig: i gjennomsnitt var 2,07 til barnet i det første og 2,73 - i det andre (av 34% mer), og dette til tross for at nivåene av reproduksjon av homoseksuelle og heterofile var nesten 5 forskjellige: henholdsvis 0,12 og 0,58 (på 383 % mindre) (Iemmola xnumx). Forskerne forklarer den uvanlig lave fruktbarheten til heterofile ved at de som kontrollgruppe skulle være så lik homoseksuelle som mulig, og derfor var de fleste ugifte. Men selv om vi tar disse ikke-representative dataene, viser det seg at for å oppnå tilstrekkelig kompensasjon vil mødre til homofile avkom trenge mer enn 7 barn ... I tillegg var det ingen signifikant forskjell i fruktbarheten til den forrige generasjonen (besteforeldre), som heller ikke er enig i oppgaven om genetisk overføre.
Forfatterne prøver å forklare de innhentede dataene og bemerker at homoseksuelle har en tendens til å overdrive antallet ikke-heterofile blant pårørende, og heterofile, tvert imot, redusere, noe som kan føre til forskjell i resultatene. De sier også at forskjeller i fruktbarhet kan forklares av fysiologiske eller atferdsmessige årsaker, som lavere abortfrekvens eller økt evne til å finne partnere. Endelig forfatterne strekat økt mors fruktbarhet forklarer mindre enn 21% av avvik i seksuell legning av menn i deres utvalg.
Dette stemmer overens med teoretiske og empiriske studier som viser at individuell opplevelse er en sterk faktor for å bestemme en persons seksuelle atferd og selvidentifisering. Det er mulig at et høyere nivå av mors homoseksualitet stammer fra kulturelle snarere enn genetisk arvede egenskaper. I mange samfunn, som Nord-Italia, tilbringer mødre mye tid med barna sine, spesielt de første årene, noe som er avgjørende for utviklingen av seksuell identitet og orientering. Dette antyder at moren og hennes familie kan være den viktigste kilden til noen oppførselsmønstre og holdninger hos barnet, inkludert trekk relatert til fremtidig seksuell preferanse og atferd. ”(Camperio-Ciani 2004).
Etter å ha gjennomført 3-studier ble forfatterne tvunget til å innrømme at dataene de mottok "De tillater oss ikke å fastslå i hvilken grad den antatte X-kromosomfaktoren fører eller til og med disponerer en mann for homo- eller bifilitet" (Ciani xnumx). Kort sagt, bidraget fra disse studiene til å forstå genesen av homoseksuell attraksjon er null.
Den største genetiske studien noensinne publisert av 30.08.2019 i en autoritativ vitenskapelig publikasjon Vitenskap, basert på et utvalg på rundt 500 tusen mennesker, fant at mer enn 99% av homoseksuell atferd bestemmes av sosiale og miljømessige faktorer. ifølge David Curtis, professor ved University of California's Institute of Genetics, "Denne studien viser tydelig at det ikke er noe som heter et homofilt gen." I den menneskelige befolkningen er det ingen slik kombinasjon av gener som vil ha en betydelig innvirkning på seksuell legning. Det er faktisk umulig å forutsi en persons seksuelle atferd etter sitt genom. ”
Del to: Hormoner?
I tillegg til genetikkens innflytelse, peker aktivister av "LGBT +" -bevegelsen på antatt intrauterin eksponering som en påstått mekanisme for den biologiske gjenstanden for homoseksuell tiltrekning. Det er underforstått at i løpet av perioden fosteret befinner seg i mors liv, virker en faktor (hormoner eller immunantistoffer) på fosteret, noe som forstyrrer den normale utviklingsprosessen, noe som videre fører til utvikling av homoseksuell tiltrekning.
For å teste hypotesen om hormonelle effekter på dannelsen av seksuell preferanse, studerer vi forholdet mellom konsentrasjonen av intrauterine hormoner på fysisk utvikling og dannelsen i tidlig barndom av atferd typisk for gutter eller typisk for jenter. Eksperimentell modellering av hormonell intrauterin ubalanse, selvfølgelig, av etiske og praktiske årsaker hos mennesker, blir ikke utført, siden hormonelle forstyrrelser fører til betydelige anatomiske og fysiologiske avvik, dette er bare mulig hos forsøksdyr7. Likevel fødes en viss prosentandel av mennesker med hormonrelatert patologi - seksuell utviklingsforstyrrelse (NDP), og i deres befolkning er det mulig å studere forholdet mellom hormonell ubalanse og atferd. Til å begynne med, bør vi kort oppgi hovedpunktene for intrauterine hormonelle effekter.
Det antas at perioder med størst reaksjon på det hormonelle miljøet oppstår under fostermodning. For eksempel er det kjent at den maksimale effekten av testosteron på det mannlige fosteret oppstår fra 8 til 24 uker, og deretter gjentas fra fødselen til omtrent tre måneder (Hines xnumx). I løpet av hele modningsperioden kommer østrogener fra morkaken og sirkulasjonssystemet til moren (Albrecht 2010). Dyreforsøk viser at det kan være mange perioder med følsomhet for forskjellige hormoner, at tilstedeværelsen av ett hormon kan påvirke handlingene til et annet hormon, og følsomheten til reseptorene for disse hormonene kan påvirke deres handlinger (Berenbaum Xnumx). Seksuell differensiering av fosteret i seg selv er et utrolig komplekst system.
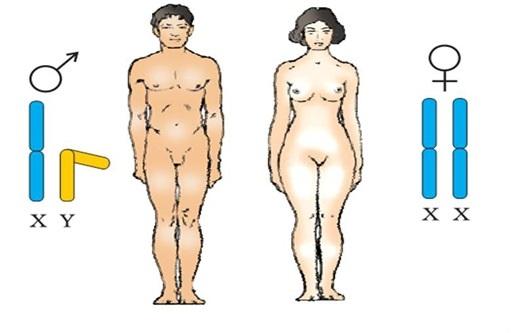
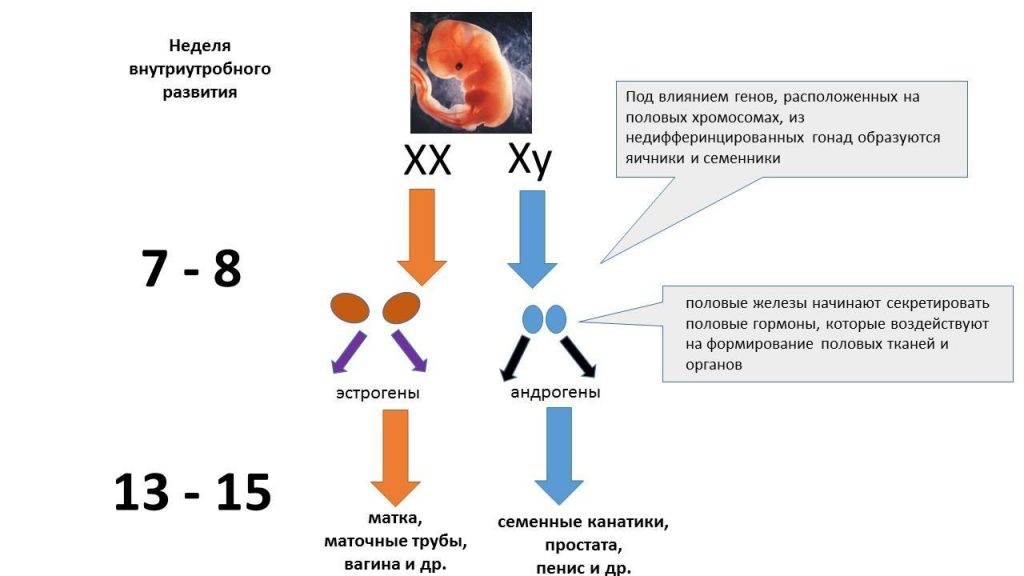
Av spesiell interesse i dette forskningsfeltet er hormoner som testosteron, dihydrotestosteron (en metabolit av testosteron og kraftigere enn testosteron), østradiol, progesteron og kortisol. Det anses som normalt hvis den hormonelle effekten på fosterets utvikling i livmoren oppstår i trinn. Helt i begynnelsen er embryoer bare forskjellige i kromosomsammensetning - XX eller XY - og deres kjønnskjertler (gonader) er de samme. Avhengig av den kromosomale kombinasjonen begynner imidlertid dannelsen av testikler i bærere av XY og eggstokkene i bærere av XX. Så snart differensieringen av gonadene er over, begynner de å produsere kjønnsspesifikke hormoner som bestemmer utviklingen og dannelsen av ytre kjønnsorganer: androgener som skilles ut av testiklene bidrar til utvikling av mannlige ytre kjønnsorganer, og fraværet av androgener og tilstedeværelsen av østrogen hos kvinner fører til utvikling av kvinnelige ytre kjønnsorganer. (Wilson 1981).
Ordningen med seksuell differensiering. Utarbeidet av V. Lysov Brudd på balansen mellom androgener og østrogener (på grunn av genetiske mutasjoner og andre påvirkninger), så vel som deres tilstedeværelse eller fravær i visse viktige perioder av fosterets utvikling, kan forårsake forstyrrelser i seksuell utvikling.
En av de mest grundig studerte forstyrrelsene i seksuell utvikling er medfødt hyperplasi av binyrebarken (VGKN), assosiert med en mutasjon av et gen som koder for et enzym som er involvert i syntesen av hormonet kortisol (Speiser 2003). Denne patologien fører til en overflod av kortisolforløpere (kortisol og androgener har en felles forløper), hvorfra androgener dannes. Som et resultat blir jenter født med ulik virilisering8 kjønnsorganer - avhengig av alvorlighetsgraden av den genetiske defekten og graden av overskudd av androgener. Alvorlige tilfeller av virilisering med utvikling av dype funksjonsfeil krever noen ganger kirurgisk inngrep. For å nøytralisere effekten av et overskudd av androgener, er hormonbehandling foreskrevet. Det ble bemerket at kvinner med HCV har større risiko for å utvikle en homoseksuell attraksjon (Speiser 2009), og de som led av HCV i en mer alvorlig form er mer sannsynlig å bli heterofile enn kvinner som har hatt sykdommen i en mildere form (Hines xnumx).
I tillegg er det nedsatt seksuell utvikling hos genetiske menn som lider av mangel på følsomhet for androgen. Hos menn med androgeninsensitivitetssyndrom produserer testiklene normalt androgen testosteron, men testosteronreseptorer fungerer ikke. Ved fødselen ser kjønnsorganene ut som kvinner, og barnet blir oppvokst som jente. Det endogene testosteronet til barnet blir omdannet til østrogen, slik at det begynner å utvikle kvinnelige sekundære seksuelle egenskaper (Hughes xnumx). Patologi oppdages bare når puberteten er nådd, når, i motsetning til etter hvert, menstruasjonen ikke begynner, og selvfølgelig er slike "kvinner" infertile, som infertilitet og "menn" med VGKN.
Det er andre seksuelle dysfunksjoner som påvirker noen genetiske menn (dvs. individer med XY-genotypen) hvis mangel på androgener er et direkte resultat av mangelen på enzymer som er involvert enten i syntesen av dihydrotestosteron fra testosteron eller i produksjonen av testosteron fra hormonforløperen. Personer med slike lidelser blir født med kjønnsdeformiteter i ulik grad (Cohen-Kettenis 2005).
I disse eksemplene er det åpenbart at homoseksuell tiltrekning og / eller valg av atferd spesifikk for det motsatte kjønn er assosiert med funksjonelle og morfologiske patologier. Slike patologier oppdages imidlertid ikke hos homoseksuelle. Antagelsen om at hormonell ubalanse på noen måte bare fører til dannelse av en homoseksuell preferanse (det vil si påvirker en atferdstrekk) og på ingen måte påvirker morfologiske og funksjonelle egenskaper, støttes ikke av empiriske observasjoner.
Det er gjort forskjellige forsøk på å identifisere eventuelle anatomiske og funksjonelle funksjoner assosiert med homoseksuell preferanse. Vurder studiene som er sitert av LGBT + -aktivister.
En studie av Simon Levey
Flere studier er utført for å studere nevrobiologiske forskjeller avhengig av seksuelle tilbøyeligheter. Den første var publikasjonen av nevrovitenskapsmannen Simon LeVay i 1991 (LeVay 1991). LeVay utførte sin forskning på resultatene av obduksjoner av avdøde mennesker. Han delte fagene inn i tre grupper - 6 "heteroseksuelle" kvinner, 19 "homoseksuelle" menn som døde av AIDS og 16 "heteroseksuelle" menn (disse parametrene er angitt i anførselstegn fordi de avdødes seksuelle preferanser i stor grad var spekulative).
I hver gruppe målte LeVey størrelsen på et spesielt område i hjernen kalt den mellomliggende kjernen til den fremre hypothalamus.9. I hypothalamus skilles flere slike kjerner fra 0.05 til 0.3 mm³ i størrelse (Byne xnumx), som er nummerert med tall: 1, 2, 3, 4. Normalt avhenger størrelsen på INAH-3 av nivået av det mannlige hormonet testosteron i kroppen: jo mer testosteron, jo større er INAH-3. LeVey uttalte at størrelsen på INAH-3 hos homofile var mye mindre enn hos menn med tiltrekning til det motsatte kjønn, mye det samme som hos kvinner. Siden strukturen i menneskekroppen bestemmes av gener, foreslo LeVey at hvis størrelsen på INAH-3 korrelerer med retningen til seksuell lyst, så "... sexlyst skyldes hjernens struktur ...", og derfor korrelerer gener med seksuell lyst.
Det skal bemerkes at LeVey viet seg fullt og helt til dette arbeidet og håpet veldig på å få nettopp et slikt resultat. Etter at hans homoseksuelle partner Richard Sherry døde av AIDS, var LeVey deprimert i noen tid (Newsweek xnumx, s. 49). Han sa til journalister etter publiseringen at han sprutet: "Jeg følte at hvis jeg ikke fant noe, ville jeg forlate vitenskapen fullstendig" (Newsweek xnumx, s. 49).
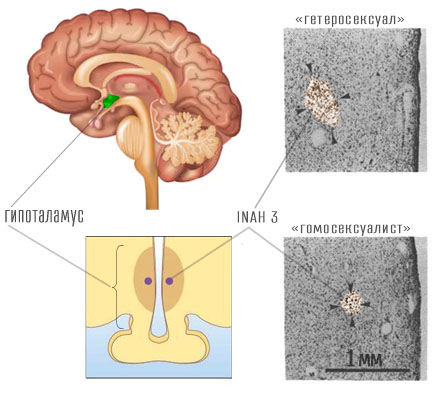
LeVey's studie hadde mange metodologiske mangler, som han selv måtte gjentatte ganger uttale, men mediene ignorerte hardnakket dem. Hva oppdaget LeVey egentlig eller ikke? Det han ikke entydig fant, er sammenhengen mellom størrelsen på INAH-3 og seksuelle tilbøyeligheter. Så langt tilbake som 1994 utsatte forsker William Byne fra New York for en alvorlig kritisk analyse av utsagnet om den genetiske årsaken til homofili (Byne xnumx): For det første er dette problemet med å velge forskningsobjekter. LeVey visste ikke nøyaktig hvilke seksuelle tilbøyeligheter menneskene han studerte i løpet av sin levetid. Det er velkjent at hos pasienter med terminal AIDS observeres lave nivåer av testosteron både på grunn av påvirkning av sykdommen og på grunn av bivirkninger av behandlingen (Gomes 2016). Fra LeVays data er det helt umulig å bestemme hvor stor INAH-3 var ved fødselen og utelukke at det kunne reduseres i løpet av livet. Alle personer som ble identifisert av LeVay som "homoseksuelle" døde av AIDS-komplikasjoner. LeVey selv tar i samme artikkel forbehold:
"... resultatene tillater oss ikke å konkludere om størrelsen på INAH 3 er en årsak eller virkning av et individs seksuelle legning, eller om størrelsen på INAH 3 og seksuell legning endrer seg gjensidig under påvirkning av noen tredje uidentifiserte variabel ..." (LeVay 1991, s. 1036).
For det andre er det ingen grunn til å si med sikkerhet at LeVey i det hele tatt oppdaget noe. Forskerne Ruth Hubbard og Elijah Wald i sin bok Destroying the Myth of Genes: How Scientists, Doctors, Employers, Insurance Companies, Educators and Human Rights Defenders Manipulerer genetisk informasjon, stilte spørsmål ved ikke bare tolkningen av LeVey's resultater, men også det faktum at noen betydningsfulle forskjeller (Hubbard xnumx, s. 95). Selv om LeVey påpekte at i gruppen av individer som han betraktet som homoseksuelle, var den gjennomsnittlige størrelsen på INAH-3 mindre enn den gjennomsnittlige størrelsen på INAH-3 i gruppen av individer som han anså som heterofile menn, følger det av resultatene hans at den maksimale og minste spredningen av verdier er perfekt det samme i begge grupper. Det er et statistisk begrep - loven om normalfordeling. Forenklet sier denne loven at det største antallet eiere av attributtet har parametrene til dette attributtet i mellomområdet, og bare et lite antall eiere har parametere med ekstrem verdi. Det vil si at av 100 personer vil 80 ha 160 - 180 vekst, 10 mindre enn 160, 10 mer enn 180 cm.
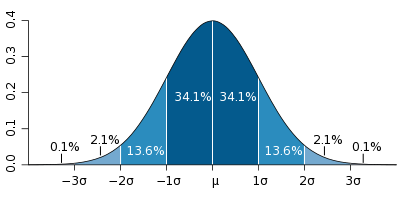
I henhold til reglene for statistiske beregninger er det umulig å sammenligne en parameter som ikke har en normal fordeling for å identifisere en statistisk signifikant forskjell mellom de to gruppene av individer. For eksempel, hvis det i en av gruppene mennesker under 160 cm ikke vil være 10%, men 40% eller 50%. I LeVays studie var INAH-3 den minste størrelsen for noen heterofile menn og de fleste homoseksuelle, og den maksimale størrelsen for noen homoseksuelle og mest heterofile menn. Det følger at for hvert enkelt individ er det helt umulig å si noe om forholdet mellom størrelsen på INAH-3 og seksuell atferd. Selv om tilstedeværelsen av noen forskjeller i strukturen i hjernen ble demonstrert overbevisende, ville deres betydning være på nivå med oppdagelsen at musklene til idrettsutøvere er større enn hos vanlige mennesker. Hvilke konklusjoner kan vi trekke på bakgrunn av dette faktum? Utvikler en person større muskler når han spiller idrett, eller gjør en medfødt disposisjon for større muskler en person til idrettsutøver?
Og for det tredje sa LeVey ikke noe om forholdet til seksuell atferd og INAH-3 hos kvinner.

I et 1994-intervju sa LeVey:
“... Det er viktig å understreke at jeg ikke har bevist at homofili er medfødt og ikke har funnet den genetiske årsaken. Jeg har ikke demonstrert at homofile blir "født på den måten" - dette er den vanligste feilen folk gjør når de tolker arbeidet mitt. Jeg fant heller ikke et “homosenter” i hjernen ... Vi vet ikke om forskjellene jeg fant ved fødselen var til stede eller dukket opp senere. Arbeidet mitt tar ikke opp spørsmålet om seksuell legning ble etablert før fødselen ... "(Nimmons xnumx).
LeVey har forbehold er veldig viktig, siden enhver spesialist innen nevrovitenskap kjenner et fenomen som nevroplastisitet - nervevevs evne til å endre funksjon og struktur i løpet av en persons liv under påvirkning av forskjellige atferdsfaktorer.
I 2000 publiserte en gruppe britiske forskere resultatene fra en hjernestudie i taxisjåfører i London (Maguire 2000). Det viste seg at for drosjesjåfører var hjerneområdet som var ansvarlig for den romlige koordinasjonen mye større enn for individer fra kontrollgruppen som ikke jobbet som drosjesjåfører, i tillegg var størrelsen på denne delen direkte avhengig av antall år brukt i en taxi (Maguire 2000). Hvis forskerne forfulgte politiske mål, kunne de ha uttalt noe som: "Disse drosjesjåførene må få utdelt høyre kjøretur, og uansett hvor de jobber, er det verdt å endre venstrekjøringen til høyre kjøretur - fordi de ble født på den måten!"

Til dags dato har en overbevisende evidensbase blitt samlet til fordel for plastisiteten i både hjernevev generelt og hypothalamus spesielt (Bains xnumx; Salg 2014; Mainardi 2013; Hatton xnumx; Teodose 1993). Hjernemorfologi endres under påvirkning av atferdsfaktorer (Kolb 1998). Hjernestrukturer, for eksempel, endres etter graviditet (Hoekzema et al. 2016)holde seg i verdensrommet (van Ombergen et al. 2018) og etter regelmessig fysisk aktivitet (Nokia et al. 2016).
Derfor, til bekreftelse av ordene som LeVey selv snakket tilbake i 1994-året, er bidraget fra hans studie av 1991-året til hypotesen om den medfødte naturen til homoseksualitet null.
En mer detaljert kritikk av LeVays arbeid, så vel som andre nevroatomiske hypoteser, gis i en gjennomgangspublikasjon i tidsskriftet Current Science (Mbugua 2003).
Replikbarheten til Levays forskning
Ingen klarte å gjenta LeVey sine resultater. I 2001-publikasjonen av året gjennomførte en gruppe forskere fra New York en lignende studie - de samme delene av hypothalamus ble sammenlignet som i LeVay-studien, men med mye mer fullstendige data og en tilstrekkelig distribusjon av de undersøkte (Byne xnumx). De fant ingen avhengighet av størrelsen på INAH-3 av homofili. Forfatterne konkluderte med at "... seksuell orientering ikke kan forutsies pålitelig basert på volumet av INAH 3 alene ..." (Byne xnumx, s. 91).
Senere ble det forsøk på å oppdage avhengigheten av seksuelle tilbøyeligheter på andre deler av hjernen. I 2002 publiserte psykologen Lasko og kollegene en studie av en annen del av hjernen - den fremre kommissuren (Lasco 2002). Det ble vist at på dette området er det ingen signifikante forskjeller avhengig av kjønn eller karakteren av seksuell lyst. Andre studier som har som mål å etablere strukturelle eller funksjonelle forskjeller mellom hjernen til heterofile og hjernen til homoseksuelle på grunn av deres iboende begrensninger, er nesten umerkelige: I 2008 ble resultatene fra noen av disse studiene oppsummert i en artikkel publisert i tidsskriftet Proceedings of the US National Academy of Sciences. (Swaab xnumx). En studie brukte for eksempel funksjonell magnetisk resonansavbildning for å måle endringer i aktivitet i hjernen når forsøkspersoner viste fotografier av menn og kvinner. Det ble funnet at å se på det kvinnelige ansiktet forsterket aktiviteten i thalamus og orbitofrontal cortex hos heterofile menn og kvinner som er homoseksuelle, mens disse homoseksuelle menn og heterofile kvinner var mer lydhør overfor mannens ansikt (Kranz 2006). At hjernen til heterofile kvinner og homoseksuelle menn spesielt reagerer på mannlige ansikter, mens hjernen til heterofile menn og homoseksuelle kvinner spesielt reagerer på kvinnelige ansikter, er vanskelig å betrakte som en stor oppdagelse, gitt etiologien til homofile tilbøyeligheter. På samme måte siterer en annen studie forskjellige reaksjoner på feromoner hos ikke-homoseksuelle menn og homoseksuelle menn (Savic 2005).
Fingerlengde
Forholdet mellom lengden på andrefingeren (pekepinnen) og fjerde fingeren (ringen) på hendene, som ofte kalles forholdet “2D: 4D”, er forskjellig for de fleste menn og kvinner. Noen bevis tyder på at dette forholdet kan avhenge av nivået av intrauterint testosteron, noe som resulterer i menn med et høyere nivå av testosteroneksponering, pekefingeren er kortere enn ringfingeren (dvs. det lave forholdet mellom 2D: 4D) og omvendt (Hönekopp 2007). Ifølge noen forskere er 2D: 4D-indeksen assosiert med homoseksuelle tilbøyeligheter. Forsøk på en eller annen måte å relatere forholdet 2D: 4D og seksuelle tilbøyeligheter er inkonsekvente og kontroversielle.
I følge en hypotese kan homoseksuelle ha et høyere forhold mellom 2D: 4D (nærmere forholdet mellom kvinner enn forholdet mellom heterofile menn), mens den andre hypotesen tvert imot antyder at hypermasculinization med prenatal testosteron kan føre til et lavere forhold homoseksuelle enn heterofile menn. En hypotese ble også fremmet om homofile tendenser hos kvinner som et resultat av hypermasculinization (lavere forhold, høyere testosteronnivå).

Flere sammenlignende studier av denne egenskapen hos homofile og ikke-homoseksuelle kvinner og menn har gitt blandede resultater. En studie publisert i tidsskriftet Nature in 2000 viste at i en prøve av 720 voksne kaliforniere, var forholdet 2D: 4D på høyre side hos kvinner med preferanser av samme kjønn betydelig mer maskuline (dvs. lavere) enn ikke-homoseksuelle kvinner, og betydelig skilte seg ikke fra forholdet hos ikke-homoseksuelle menn (Williams 2000). Denne studien avdekket heller ingen signifikant forskjell mellom gjennomsnittet av 2D: 4D mellom homofile menn og homofile. Samme år viste en annen studie som brukte en relativt liten prøve av homoseksuelle og ikke-homoseksuelle menn fra Storbritannia en lavere verdi av 2D: 4D (dvs. mer maskuline) blant homoseksuelle (Robinson 2000). I et 2003-år fant en studie av et utvalg av londonere at homoseksuelle hadde en lavere hastighet på 2D: 4D sammenlignet med ikke-homoseksuelle menn (Rahman xnumx), mens to andre studier av prøver fra California og Texas viste høyere verdier av 2D: 4D for homoseksuelle (Lippa xnumx; McFadden 2002). I 2003 ble det foretatt en sammenlignende studie av syv par monozygotiske tvillingkvinner, i alle par hadde en av tvillingkvinnene homoseksuelle preferanser, og fem par monozygotiske tvillingkvinner der begge søstrene hadde samme kjønnspreferanser (Hall 2003). I par tvillinger med forskjellige typer seksuell tiltrekning, hos individer som identifiserer seg som homoseksuelle, var forholdet 2D: 4D betydelig lavere enn for tvillingene, mens de konkordante tvillingene ikke fant noen forskjell. Forfatterne konkluderte med at dette resultatet indikerer at "det lave forholdet mellom 2D: 4D er resultatet av forskjeller i prenatal miljø." Og til slutt, i 2005-året, som et resultat av en studie av forholdet 2D: 4D i et østerriksk utvalg av 95 homoseksuelle menn og 79 ikke-homofile menn, ble det funnet at indikatorene 2D: 4D hos ikke-homofile menn ikke skilte seg nevneverdig fra de hos homofile menn (Voracek 2005). Etter å ha gjennomgått flere studier av denne egenskapen, konkluderer forfatterne at "mer data er nødvendig for å kunne konkludere med selvtillit om det er en sammenheng mellom forholdet 2D: 4D og arten av seksuell lyst hos menn, underlagt etniske forskjeller."
Øyeblikk
I 2003 kunngjorde en gruppe engelske forskere at de hadde funnet "nye overbevisende bevis på at seksuell lyst skyldes egenskapene til den menneskelige hjernen" (Rahman xnumx). Katsi Rahman og medforfattere sa at de fant en forskjell i reaksjonshastighet - blinkende øyne - som svar på høye lyder. Forfatterne fant at kvinner har mindre såkalte "Pre-puls inhibition" (PPI) - en reduksjon i kroppens motoriske respons på stimuli, i nærvær av en svak foreløpig stimulus10... Det vil si at kvinner blinket raskere enn menn, og kvinner med samme kjønn preferanse blinket langsommere enn ikke-homofile kvinner. Det skal bemerkes at for det første gjennomførte forfatterne en studie i en liten gruppe fag, og for det andre fant de ingen forskjeller mellom homofile menn og ikke-homofile menn. Uansett bestemte forfatterne at resultatene deres beviser at homofili er et medfødt fenomen. Ikke desto mindre tok forskerne likevel flere forbehold: De bemerket at spørsmålet om forskjellene som er funnet skyldes spesifisiteten til seksuell tiltrekning eller er resultatet av en viss seksuell oppførsel, er fortsatt ikke løst. De påpekte: "... de nevroanatomiske og nevrofysiologiske variasjonene mellom heterofile og homofile kan skyldes enten biologiske faktorer eller innflytelse fra læring ...". Dr. Halstead Harrison fra University of Washington analyserte studien og bemerket en så viktig mangel som den lille størrelsen på testgruppene (14 homofile kvinner og 15 heterofile kvinner, 15 homofile menn og 15 heterofile menn). Harrison konkluderte: "Rahman et al. Har ikke gitt avgjørende bevis for å støtte konklusjonen om at homofile kvinner viser PPI-parametere som ligner på menn."Harrison xnumx). Harrison stilte også spørsmål ved metodenes statistiske tilstrekkelighet.
Tvillingstudiene diskutert over kan belyse graden av innflytelse av mors hormoner, siden identiske og identiske tvillinger under intrauterin utvikling utvikler sin effekt på samme måte. Svake indikatorer på samstemthet i tvillingstudier indikerer at prenatale hormoner som genetiske faktorer ikke spiller en avgjørende rolle i dannelsen av seksuell lyst. Andre forsøk på å finne hormonelle faktorer som vesentlig påvirker seksuell lyst, har også vært entydige, og betydningen av resultatene deres er ennå ikke forstått.
Effektene av mors stress
I 1983 gjennomførte Gunther Dörner et al en studie for å etablere en kobling mellom mors stress under graviditet og den påfølgende seksuelle identiteten til barna deres. De intervjuet to hundre mennesker om hendelser som kan forårsake stress hos mødrene under graviditet - det vil si den intrauterine utviklingen til respondentene selv (Dörner 1983). Mange av hendelsene var relatert til kjølvannet av 2. verdenskrig. Av mennene som rapporterte at mødrene deres opplevde moderat til alvorlig stress under graviditeten, var 65% homoseksuelle, 25% var bifile og 10% var heterofile. I senere studier ble det imidlertid observert enten mye mindre korrelasjoner eller fraværet av signifikante korrelasjoner (Ellis 1988). I 2002, etter å ha utført en prospektiv studie av forholdet mellom sexlyst og prenatal stress i andre og tredje trimester, fant Hines og kolleger at mors stress under graviditet "bare var litt relatert" til den typiske mannlige oppførselen til deres døtre i en alder av 42 måneder " og ikke noe forhold til ”sønnenes typiske feminine oppførsel (Hines xnumx).
Del tre: Immunforstyrrelser?
Big Brother-effekten
“Effekten av den eldre broren” (ESB) eller “effekten av rekkefølgen på brødre”11 - dette begrepet ble foreslått av kanadisk-amerikanske forskere ved navn Ray Blanchard og Anthony Bogert - det er at ifølge noen observasjoner, sammenlignet med normale heterofile menn, har homofile pedofiler, homofile og voldtektsmenn flere eldre brødre, men ikke eldre søstre (Blanchard 1996; Bogaert 1997; Blanchard 1998; Lalumiere 1998; Blanchard 2000; Cote xnumx; MacCulloch 2004; Blanchard 2018).

For øyeblikket gjenstår det en åpen diskusjon om hvorvidt (1) om en ESB virkelig eksisterer, og (2) om den eksisterer, om den har en biologisk eller sosial årsak (Zietsch 2018; Gavrilets 2017; Whitehead 2018).
Til tross for de motstridende resultatene innen ESB og dets årsaker, aksepterte noen forskere og offentlige personer, som prøvde å finne biologiske begrunnelser for homofili, så klart den biologiske forklaringen til ESB at de helt utelukket alle andre mulige forklaringer (påvirkning av oppdragelse, etc. .).
⚡️2023 tillegg:
Vilsmeier JK, Kossmeier M, Voracek M, Tran US. 2023. Den broderlige fødselsrekkefølgeeffekten som en statistisk artefakt: konvergent bevis fra sannsynlighetsregning, simulerte data og multivers metaanalyse. Peer J 11:e15623 https://doi.org/10.7717/peerj.15623
Forskere fra Institutt for psykologi ved Universitetet i Wien utførte matematisk behandling av data om storebror-effekten. De konkluderte med at, når de ble riktig analysert, er den spesifikke assosiasjonen mellom antall eldre brødre og homoseksuell legning liten, heterogen i størrelsesorden og tilsynelatende ikke spesifikk for menn. Dessuten eksisterende vitenskapelig bevis overdrevet på grunn av effektene av små studier.
Ulemper ved ESB-hypotesen
ESB er ikke et ubetinget aksiom, faktum om eksistensen er gjenstand for pågående vitenskapelig diskusjon av mange grunner.
For det første blir denne effekten ikke påvist i alle studier. Brendan P. Zietsch bemerket at tilhengere av ESB-hypotesen i sine analyser bare inkluderer resultatene fra publiserte studier som er i samsvar med deres ideer, og ignorerer studier, nyhetsbrev, avhandlinger, presentasjoner på konferanser der ESB ikke blir oppdaget (Zietsch 2018). Dette problemet er spesielt viktig, gitt at ESB ikke ble bekreftet i seks av sju korrekt like sannsynlighetsprøver (Bearman 2002; Bogaert 2005, 2010; Francis xnumx; Frisch xnumx; Zietsch 2012). LGBT + -aktivisten, nevnt over, av Simon LeVay-bevegelsen, gir i sitt arbeid også en oversikt over studier der ESB ikke ble oppdaget (LeVay 2016).
For det andre er de studiene der ESB ble påvist basert på en tvilsom prøvetakingsmetodikk. Støttespillere av ESB-hypotesen bruker slike kriterier for populasjonsanalyse som fører til ekskludering av alle tilgjengelige sannsynlighetsprøver (dvs. de prøvene som er tilfeldig valgt i forhold til den studerte uavhengige variabelen - seksuell tiltrekning i dette tilfellet). Dette betyr at metaanalysen bare inkluderer de prøvene der andelen homoseksuelle ikke ligner andelen homoseksuelle i den generelle befolkningen (for eksempel prøver fra Blanchard-analysen av året 2018 i gjennomsnitt inneholder 51% av homoseksuelle, mens i den generelle befolkningen av dem, i henhold til forskjellige kilder er maksimum 2 - 3%). Når det gjelder slike ikke-tilfeldige prøver øker risikoen for å velge homoseksuelle og heterofile grupper, noe som ikke bare skiller seg i prediktorvariabler. Blanchard 1-tabellen 2018 viser at de fleste prøvene som er inkludert i metaanalysen er hentet fra ekstremt urepresentative populasjoner: sexkriminelle, transpersoner, pedofile, psykopater, etc. Det er bemerkelsesverdig at ingen av disse problemene med utvalg av utvalg ble diskutert i artikkelen. Tvert imot ble Blanchards inkluderingskriterier anvendt på en slik måte at det ekskluderte store studier med sannsynlighetsprøver (der ESB ikke ble bekreftet). Den store heterogeniteten i størrelsen på effekten mellom de individuelle studiene i metaanalysen viser at det faktum hvordan gruppene er valgt ut til studien, har stor innflytelse på ESB. Dette øker sannsynligheten for at funksjonene i prøven skaper ESB, spesielt med tanke på at store sannsynlighetsprøver ikke viser ESB i det hele tatt.
For det tredje er et annet metodologisk problem at analysemetodene for å finne ESB-er virker partiske og rettet mot å oppdage ønsket effekt. Noen forskere brukte for eksempel en enveis statistisk test for å måle effekten (f.eks. Bogaert 2005; Poasa 2004; Purcell 2000) eller tolket resultatene fra andre forskere som faktisk ikke oppdaget ESB som betydningsfulle, og sa at enveis-tester burde vært brukt (Blanchard 2015) - selv om det er kjent at enveis tester bare kan brukes i svært sjeldne tilfeller som ikke passer til metaanalysen (Lombardi xnumx). Forsker Bartlett skriver følgende:
“... Gitt den relative mangelen på homofile menn i befolkningen, er det vanskelig å finne balanserte grupper av homofile og heterofile menn for studien. Prøvetakingen av homofile og heterofile fra befolkninger med forskjellige familiestørrelser utgjør et problem i måling av ESB. Sannsynligheten for at studien vil finne en falsk effekt hos alle slags søsken, ikke bare eldre søsken, øker hvis homofile fra større familier blir valgt, mens effekten forsvinner hvis heterofile menn fra større familier blir valgt ut i utvalget. ... "(Bartlett xnumx).
For det fjerde er ESB utelukkende basert på resultatene fra korrelasjonsanalyse. Påvisningen av faktiske korrelasjoner er identisk med deteksjonen av årsaken som skaper denne korrelasjonen. Eventuelle korrelasjoner trenger også en mekanistisk forklaring av hva som ikke ble oppfylt (Gavrilets 2017).

For det femte er ESB ikke universell. ESB er ikke i stand til å forklare homofili hos menn som ikke har eldre brødre, og er heller ikke i stand til å forklare mangelen på homoseksuell tiltrekning hos yngre brødre som har en eldre bror til homofili, kan ikke forklare misforholdet til seksuelle preferanser blant tvillingbrødrene12. ESB forekommer ikke hos bifile menn. Biseksuell tiltrekning kan forstås som en seksuell tiltrekning både motsatt og ens eget kjønn. Derfor, innenfor rammen av ESB-paradigmet, bør biseksuelle menn ha mindre ESB enn homoseksuelle menn, men mer enn heterofile menn. Imidlertid i studien Bogaert (2006) ESB var den samme for bifile og homoseksuelle individer. McConaghy og kolleger (2006) gjennomførte en ESB-studie i “hovedsakelig heterofile individer” (individer med en liten tiltrekning av samme kjønn) sammenlignet med en kontrollgruppe av eksepsjonelle heterofile. ESB er observert for både menn og kvinner. I tillegg ble effekten av storesøsteren også observert hos menn, selv om den var mindre sterk. Ifølge forfatterne indikerer resultatene at biologiske årsaker til ESB er mindre sannsynlige enn sosiale. Det er anslått at ESB-hypotesen bare forklarer 17% av det totale antall tilfeller av homoseksuell tiltrekning og bare hos menn (Kantor xnumx). ESB forklarer ikke homoseksuelle preferanser hos kvinner. Tilhengere av ESB-hypotesen har mange ganger prøvd å finne denne effekten hos kvinner med homoseksuelle preferanser, men uten resultater (Blanchard 2004).
For det sjette fungerer ESB ikke i virkelige kultur-etniske prediktive modeller. Forutsatt at det finnes en ESB, i henhold til dens paradigme, kan man forutsi (modellen iht Bogaert 2004) at en stor utbredelse av menn med homoseksuelle preferanser blir observert i: (a) religiøse familier, der sannsynligheten for et stort antall barn er høyere; (c) østlige og muslimske kulturer, tradisjonelt preget av store familier; og lavere utbredelse - i vestlige samfunn med høy levestandard, der fødselsraten er betydelig dårligere enn østlige samfunn (Caldwell 1997). En lignende trend tilsvarer mildt sagt ikke virkeligheten.
ESB-hypoteser
Det er flere antagelser som forklarer ESB funnet i noen studier (James xnumx), blant dem kan to hovedtrekk skilles: (1) biologisk prenatal eksponering (mors immuniseringshypotese) og (2) sosialt psykologisk postnatal (eksponering for miljøforhold). Nedenfor vil vi analysere begge forutsetningene.
Immuniseringshypotese om mor
Som det biologiske grunnlaget for ESB la Blanchard og Bogert frem hypotesen om immunkonflikt i moren, som var at det kvinnelige immunsystemet visstnok er i stand til å produsere antistoffer mot visse "mannlige antigener" i det mannlige fosteret, og antas at lignende antistoffer akkumuleres med hvert påfølgende svangerskap av det mannlige fosteret, øke risikoen for intrauterin immunskade for hver påfølgende gutt (Blanchard 1996). Hypotesen om immunkonflikt i mor prøver å forklare utviklingen av guttens homoseksuelle preferanser på analogi med Rh-konflikt graviditet (Bogaert 2011).
Rhesus-konflikt graviditet er en patologisk tilstand forårsaket av tilstedeværelsen i fosteret av et gen som koder for et spesifikt protein på blodceller og fraværet av et slikt gen i moren (det vil si at moren i dette eksemplet er Rh-negativ og fosteret er Rh-positiv). I løpet av det første svangerskapet til en Rh-negativ mor med et Rh-positivt foster, trengs cellene i fosteret inn i blodstrømmen til moren og forårsaker en immunreaksjon - dannelse av antistoffer mot blodceller. I påfølgende graviditeter hos denne moren med et Rh-positivt foster, vil antistoffer fra mors blodomløp trenge inn i fosterets blod og ødelegge dets røde blodlegemer, forårsake hemolyse og gulhet ved fødselen. Det er derfor fødselslege-gynekologer kontrollerer Rh-statusen til den gravide moren og faren til barnet.
Blanchard- og Bogert-hypotesen er basert på de samme prinsippene som Rh-konflikt graviditet. I dette tilfellet er faktoren som forårsaker dannelse av antistoffer (Rh-positivitet i eksemplet ovenfor) tilstedeværelsen av et lekekromosom, det vil si fosterets mannlige kjønn. Y-kromosomet koder for dannelse av proteiner og hormoner som er til stede i det mannlige fosteret (men ikke hos kvinnelige!) Allerede i de tidligste stadiene av embryogenese. I henhold til hypotesen diskutert, kommer partikler av fostervev som bærer det "mannlige antigenet" inn i blodstrømmen til moren og forårsaker dannelse av antistoffer, som visstnok, under påfølgende svangerskap av det mannlige fosteret, krysser blod-hjerne-barrieren, trenger inn i fosterhjernen og angriper spesifikke nerveceller som inneholder det "mannlige antigenet". ", Angivelig forhindrer utviklingen av den embryonale hjernen" av den mannlige typen ", som et resultat av at gutten er født med en" kvinnelig hjerne "og visstnok blir homoseksuell eller transkjønn. Mors immunreaktivitet øker med hvert nytt svangerskap av det mannlige fosteret, derfor øker sannsynligheten for avvik angivelig med hver eldre bror.
I henhold til hypotesen fra Blanchard og Bogert er bekreftelse av intrauterin immunskade en reduksjon i kroppsvekt ved fødselen hos homoseksuelle menn som har eldre brødre.
Ulemper ved immuniseringshypotesen om mor
William H. James (2004) kritisk undersøkt de grunnleggende elementene i hypotesen om immun immunkonflikt i moren.
For det første antakelsen om at moren under graviditet blir immunisert bare med spesifikke antigener fra det mannlige fosteret, men ikke det kvinnelige - for å si det mildt, er det tvilsomt. Mødre kan utvikle immunreaksjoner mot fosteret, både hann og kvinne, det vil si ikke "mannlige antigener", men spesifikke fader har immunreaktivitet i disse tilfellene, og slike patologier er godt studert (Dankers xnumx). Tre slike reaksjoner er vanligst: (a) den nevnte RCH, der føtal røde blodlegemer er påvirket, som har en positiv Rh-faktor på overflaten, frekvens 10 - 20%; (b) alloimmun trombocytopeni hos nyfødte som påvirker blodplater, frekvens 4% eller 12%, hvis det også tas hensyn til asymptomatiske former (Turner 2005); nøytropeni hos nyfødte, påvirker nøytrofiler, frekvens 4% (Han 2006). I alle disse tilfellene er antigener individuelle faderlig, ikke vanlig hann. De utvikler seg til etterfølgende barn av alle kjønn fra samme far. De påvirker blodkomponenter (og ikke visse organer og vev) under kontakten med fosterets blod (navlestreng, morkake, etc.) med mors immunsystem (på grunn av traumer til de ytre kjønnsorganene, livmorens indre overflate, etc.) under fødsel.
Moroalloimmune antistoffer trenger visstnok trenge inn i morsmelken, som alle andre antistoffer (Gasparoni xnumx), for eksempel alloimmune mors antistoffer mot Rh-faktoren, som trenger inn i morsmelken, kan føre til hemolytisk sykdom hos det nyfødte (Øl 1975). Tilsvarende kan det antas at melk som inneholder hypotetiske antistoffer mot "mannlige antigener", tolereres dårlig av senere brødre, noe som vil føre til problemer med amming og tidlig opphør, samt allergisk kolitt. En gjennomgang av medisinsk litteratur gir imidlertid et helt motsatt bilde: Fødselsrekkefølgen er ikke relatert til ammingens varighet eller korrelerer generelt positivt med den (Martin 2002). Hyppigheten av allergisk kolitt hos nyfødte varierer fra 0,01% til 7,5% (Hildebrand xnumx; Pumberger xnumx; Xanthakos 2005), mens nyfødte av begge kjønn blir berørt. Også inkludert i denne statistikken er reaksjoner på kumelk.
Vi gjentar at fra et evolusjonært synspunkt er den intrauterine immunogenisiteten til det mannlige fosteret absurd for moren. Humant fylogenese som et pattedyr varer i mange millioner år. Hvorfor i så lang tid i menneskekroppen ikke har utviklet effektive måter å forhindre så kostbare fra synspunktet om evolusjon av immunresponser? Hypotetiske immunreaksjoner av kvinnekroppen under den evolusjonært så rutinemessige og uunngåelige prosessen for en sunn kvinnelig kropp som graviditet med et mannlig foster, som utgjør 50% av alle svangerskap, ville føre til betydelig seksuell ubalanse og evolusjonsproblemer. Filogenese fører alltid til seleksjon og bevaring av de mest optimale egenskapene for arten. For eksempel er det signifikante bevis for at valget av en mannlig partner er assosiert med et hovedhistokompatibilitetskompleks (GCS) (Chaix 2008; Millinski 2006; Wedekind xnumx), det vil si på fylogenetisk nivå, er artsprosesser maksimalt rettet mot å øke mangfoldet på grunnlag av GCS og øke levedyktigheten til avkom (Williams 2012; Guleria 2007).
Til forsvar for sin teori gir Bogert som eksempel en slik patologisk immunrespons som Rh-conflict graviditet (RCH) (Bogaert 2011), noe som fører til hemolytisk sykdom hos det nyfødte - angivelig dette fenomenet (med risiko er omtrent 15% av befolkningen (Izetbegovic 2013)) forsvant ikke under evolusjonen. Det må imidlertid tas i betraktning at hyppigheten av FC i menneskehetens fortid som art var betydelig lavere. På det nåværende stadiet observeres en slik evolusjonær faktor som forvirring av menneskeheten, derfor virker det ikke paradoksalt at de naturlige mekanismene for å blokkere Rhesuskonflikten ennå ikke har utviklet seg. Med utviklingen av transplantologi har menneskeheten møtt en slik faktor som tidligere var fraværende som immunavstøtningsreaksjonene (i nesten 100% av mottakerne), er det ikke overraskende at mennesker ikke har noen naturlig mekanisme for undertrykkelse. Når det gjelder RCH og transplantasjonsavstøtningsreaksjoner for en person som art, har det ikke gått mye tid for utvikling av kompensasjonsmekanismer13. På den annen side ville det stabile vedlikeholdet av immunkompatibiliteten til mødre med 50% av avkommet være paradoksalt.
Generelt virker det tvilsomt at det er visse strukturer eller stoffer i det mannlige fosteret som kun har antigene egenskaper som er spesifikke for hannen. Gratis testosteron, et kjønnshormonbindende globulin eller cellemembran androgenreseptor, er ikke immunreaktivt for moren, fordi alle av dem også er til stede i kvinnekroppen.
For det andre antakelsen om at spesifikke moderantistoffer selektivt skader hjernen til det mannlige fosteret (noe som fører til det "feminisering"), men samtidig forstyrrer de ikke andre hjernefunksjoner og påvirker ikke testiklene (inneholder mye mer produkter av Y-kromosomgener) ) - er mildt sagt kontroversiell.
Hvis det faktisk skjedde en immunreaksjon mot "mannlige antigener", ville hypotetiske morsantistoffer først og fremst eller hovedsakelig eller i det minste samtidig påvirke testiklene, som inneholder mye mer "mannlig antigen" enn hjernen. Mange mannspesifikke gener er kjent (dvs. lokalisert på Y-kromosomet) (Ginalksi xnumx). Uttrykket av disse genene - det vil si lesing av informasjon og syntesen av proteiner og strukturer - forekommer ikke bare og ikke så mye i hjernen, men hovedsakelig i testiklene, som bør være det primære målet for det "anti-mannlige" spesifikke immunangrepet, og ikke hjernen. (Ginalksi xnumx). Hos homoseksuelle menn vil en økt forekomst av testikkelpatologier kunne observeres: hypospadi, kryptorkidisme, testikkelkreft, etc., men noen forbindelse av testikkelforstyrrelser med homofili eller ESB ble imidlertid ikke funnet (Pierik xnumx; Flannery xnumx). Dessuten er det interessant å merke seg at menn med hypospadi, til tross for lave testosteronnivåer under prenatal utvikling, har litt høyere nivåer av psykologisk maskulinitet (Sandberg 1995). Det kan også forventes at hos individer med homoseksuell tiltrekning, pubertet vil oppstå senere på grunn av immun testikkel lesjoner, men store studier avslørte imidlertid ikke forskjeller i pubertetsalderen avhengig av seksuelle preferanser (Savin-Williams 2006).
I tillegg vil inntreden av hypotetiske morsantistoffer gjennom blodomløpet inn i fosterhjerne være umulig på grunn av blod-hjerne-barrieren (BBB), som er dannet allerede den 4-th uken av svangerskapet (Zusman 2004). Slike antistoffer ville være i stand til å overvinne BBB bare med alvorlige patologier fra sistnevnte - med brudd på beskyttende funksjoner, noe som vil føre til betydelig nevrologisk skade på hjernen. Imidlertid, hvis fosterets BBB er i normal tilstand, fører til og med brudd på morens immunsystem ikke til nevrologiske patologier hos det nyfødte - BBB forhindrer antistoffer. I en stor studie som dekket 17 283 par mødre med barn, ble det ikke funnet noen sammenheng mellom økt mors immunreaktivitet og cerebral parese, psykisk utviklingshemning, kramper osv. (Flannery xnumx).
Hypotesen om at hypotetiske antistoffer skader hjernen på en slik måte at de forårsaker feminisering er uholdbar. I fasen av embryogenese er anatomiske kjønnsforskjeller i hjernen svakt uttrykt, og den endelige morfofunksjonelle dannelsen av hjernen, ifølge kjønn, oppstår i puberteten, når en hypotetisk immuneffekt er umulig (Lenroot 2007; Paus xnumx). Selve ideen om tilstedeværelsen i hjernen til et embryo av en nevral organisasjon som er karakteristisk for et bestemt kjønn, er veldig tvilsomt og har aldri blitt overbevisende demonstrert (Lauterbach 2001; Nunez 2003). MR-skanninger viste bare ubetydelige statistiske snarere enn dikotome forskjeller i hjernestrukturen til nyfødte, med betydelige samsvar mellom kjønnene (Zanin xnumx; Mitter 2015).
I henhold til hypotesen, bør vi forvente at homoseksuelle med eldre brødre, som har en "feminisert" hjerne, alltid vil tilhøre fenotypen med typisk kvinnelige interesser og atferd, siden det er ekstremt spekulativt å tro at "demaskuliniseringen" av hjernen bare vil påvirke guttens seksuelle preferanser, men vil omgå andre spesifikke mannlige egenskaper. Det skal bemerkes at i noen studier er det samme kjønnsattraksjon hos voksne som korrelerer med mer "kvinnelige" hjernestrukturer, men hjerneutvikling, når det gjelder størrelse og funksjon, skjer hovedsakelig etter fødselen, og derfor er slike strukturer, ifølge forfatterne selv, resultatet av postnatal erfaring, ikke fødselsfaktorer. Forskning av Bogaert et al. (2003; 2005); Kishida et al. (2015); Semenyna et al. (2017) avslørte ikke sammenhenger mellom ESB og alvorlighetsgraden av feminine tegn hos menn.
For det tredje er forholdet mellom en hypotetisk intrauterin immunlesjon, antall eldre brødre, homoseksuell tiltrekning og vekttap ved fødselen tvilsom.
Som bevis på et generelt immunangrep siterer talsmenn for ESB-hypotesen og immunskadene data om at menn med eldre brødre hadde lavere fødselsvekt (Blanchard 2001). Nedgangen i kroppsvekt ved fødselen hos gutter som har eldre brødre, i Blanchards studier var omtrent 170 gram (5% av kroppsvekten) (Blanchard 2001). I henhold til hypotesen som diskuteres, bør en lignende reduksjon observeres for gutter med homoseksuell preferanse som har eldre brødre, og bør ikke observeres hos jenter. Dette er imidlertid ikke slik - i en norsk studie som undersøkte den hypotetiske sammenhengen mellom immunsvar og vekttap ved fødselen, ble 181 000 fødselsstilfeller studert, og vekttap ved fødselen ble observert hos både jenter og gutter (Magnus 1985). Videre ble den hypotetiske "storebror-effekten" notert for begge kjønn og var ekstremt lav - 0,6%, uttrykt i forskjellen på 20 ± 4,5 gram med hensyn til standard fødselsvekt i 3 500 gram (Magnus 1985).
I følge disse dataene virker immunfaktorers rolle generelt i å redusere kroppsvekten tvilsom. Det er bemerkelsesverdig at Magnus og kolleger i deres studie også studerte effekten av fosterantigener på vekten av nyfødte - i dette tilfellet ble det antydet at hvis vekttap er forårsaket av immunantistoffer mot faderlige antigener, vil det bli notert hos både gutter og jenter .. Magnus og kolleger studerte kroppsmassen til barn av begge kjønn ved fødselen hos mødre som inngikk et nytt ekteskap og fødte nye barn - hvis vekttapet skyldtes immunrespons, burde fødselsvekten hos barn til en annen mann ha vært å gå tilbake til standardindikatorene, siden den andre faren er en bærer av nye antigener og en progressiv immunprosess er nødvendig for akkumulering av immunantistoffer (flere graviditeter) (Magnus 1985). Imidlertid forble kroppsvekten ved fødselen av barn fra en annen far redusert, og forfatterne konkluderte med at forholdet mellom eventuelle immunprosesser og en reduksjon i kroppsvekt ved fødselen ikke er bekreftet i deres utvalg (Magnus 1985).
Årsaken til vekttap ved fødselen kan være: (a) prematuritet; (b) placental insuffisiens; (c) moroimmune sykdommer, for eksempel systemisk lupus erythematosus (kombinert med en rekke medfødte patologier ved fødselen); (d) et kompleks av patologier assosiert med testikkelforstyrrelser. Ingen av de ovennevnte er blitt kjent for homofile menn som har eldre brødre.
Forholdet mellom vekttap ved fødselen og immunresponsene er ikke avklart og er fortsatt et veldig spekulativt spørsmål. I følge James (2006) en markant reduksjon i kroppsvekt ved fødselen kan skyldes påvirkning av testosteron (Manikkam 2004). I tillegg er økte nivåer av testosteron i kvinnekroppen assosiert med økt sannsynlighet for å føde en gutt (James xnumx; James 2004b). Blanchard refererte til en studie når han utviklet hypotesen sin om kvaliteten på bevisene som støtter den Gualtieri og Hicks (1985)som uttalte at den seksuelle andelen barn som ble født skifter mot det kvinnelige kjønn avhengig av antall barn (med andre ord, jo flere barn ble født i familien, desto mindre sannsynlig var gutten å bli født). Imidlertid var det en tolkningsfeil i denne studien (se James xnumx, s. 52; James xnumx). I kontrast, de to største studiene: en analyse av 4 millioner fødsler i Frankrike (James xnumx) og 150 tusen fødsler i USA (Ben-porath xnumx) avslørte at sannsynligheten for å føde en gutt øker med en økning i antall eldre brødre og avtar med en økning i antall eldre søstre, noe som strider mot ESB. Biggar et al. (1999) Basert på disse dataene, utførte vi en statistisk analyse av 1,4 på en million fødsler og fant ut at sannsynligheten for å få en gutt øker med en økning i antall eldre brødre.
For det fjerde er antakelsen om at den førstefødte gutten i familien ikke skal ha homoseksuelle preferanser, og følgelig risikoen for deres utvikling øker med økningen i antall eldre brødre, er mildt sagt spekulativ.
Ikke alle homofile menn har eldre brødre, derimot er noen eldre brødre eller bare gutter i familien homoseksuelle. Tilhengere av hypotesen fremmet et motargument om at mødrene til slike menn angivelig hadde spontane aborter av mannlige foster før fødselen deres, noe som utløste immuniseringsprosessen. Forekomsten av par med spontanaborter er 1%; i omtrent halvparten av disse tilfellene har fosteret en normal karyotype, det vil si at det kan antas at halvparten av spontane aborter er forårsaket av immunreaksjoner (Lee 2000). Undersøkelser om kjønnsforholdet til embryoer som dør som et resultat av spontan abort viser imidlertid at mer enn halvparten var kvinnelig: mann / kvinneforholdet er 0,76 (Eiben xnumx) 0,71 (Eiben xnumx) 1,03 (Være xnumx); 0,77 (Smith 1998) 0,77 (Evdokimova 2000) 0,83 (Morikawa xnumx) 0,35 (Halder 2006) 0,09 (Kano xnumx).
På den annen side, i henhold til immunhypotesen, skal hjernen til hvert mannlig foster i livmoren angripes med økende intensitet i alle påfølgende graviditeter, det vil si gjennomgå mer og mer "feminisering", men dette er ikke slik. Ikke alle yngre brødre til en homofil mann har homoseksuelle preferanser. Interessant nok er de yngre brødrene til menn med et brudd på kjønnsidentitet - hvis hjerne, ifølge Blanchards hypotese, skal gjennomgå "feminisering" - utvikles normalt (Grønt antall).

Kilde: Michael Ochs Archives, Getty Images
I følge hypotesen kan det også forventes at brødre født senere vil lide av mange fysiske problemer på grunn av økte immunologiske angrep fra moren, men det motsatte er sant: den senere fødselsrekkefølgen er hovedsakelig forbundet med en forbedring snarere enn en forverring helse (Juntunen xnumx; Cardwell xnumx; Sorenson 2005; Richiardi xnumx).
Hypotese om sosial innvirkning forklaring av ESB
Forfatterne av mødreimmuniseringshypotesen bemerket selv:
“… Det er selvfølgelig andre mulige forklaringer på storebroreffekten i tillegg til hypotesen om mors immunrespons. Den mest populære konkurrerende hypotesen er at seksuell interaksjon med voksne menn øker sannsynligheten for at en gutt utvikler homofil tiltrekning, og at guttens sjanser for å delta i slike interaksjoner øker proporsjonalt med antallet hans, antallet eldre brødre ... "(Ellis 2001).
Velvære og kolleger (1994, s. 204 - 206) fant at menn som gikk på guttenes internatskoler var mer sannsynlig å rapportere om homoseksuell opplevelse i løpet av livet enn menn som ikke gikk på slike skoler, men det var ingen forskjell i andelen individer som rapporterer om homoseksuelle opplevelser senere i livet. ” Blanchard (Ellis 2001) henvist til publisering Velvære og kolleger (1994) som bevis på at den sosiale hypotesen ikke er relevant. De tolket imidlertid disse dataene på en særegen måte. Wellings på side 206 gir en graf som viser at omtrent 1,5% av 7925 menn som ikke har gått på internat rapporterte mer enn en homoseksuell kontakt de siste 5 årene, og 2% av 412 menn som gikk på skolen internat. Det er klart at disse dataene (uforholdsmessige størrelser på grupper) snakker mer sannsynlig til fordel for den sosiale hypotesen. Vurder andre studier i forbindelse med sosial teori.
Blanchard selv antydet at blant mannlige pedofiler var omtrent 25% homofile pedofiler (Blanchard 2000b). Dette er omtrent ti ganger andelen homofile blant menn hvis seksuelle interesser er rettet mot voksne menn. Det er blitt antydet at blant menn har homofili og pedofili en vanlig årsak, og denne årsaken er seksuelle (eller kvasi-seksuelle) opplevelser i en tidlig alder (James 2004). I følge denne ideen vil tidlig homofil erfaring undertrykke dannelsen av seksuell interesse for det motsatte kjønn i voksen alder. Rimafedi (1992) fant at usikkerhet om egne seksuelle preferanser hos ungdommer avtar med alderen: disse forfatterne antyder at seksuell identitet utvikler seg i ungdomsårene og påvirkes av seksuell erfaring.
Dessuten observeres hyppigere tilfeller av seksuell vold i barndommen blant homofile menn enn blant heterofile menn (Paul 2001; Finkelhor xnumx, 1984); det var en betydelig sammenheng mellom et mannlig seksuelt overgrep og seksuell krenkelse (Glasser 2001); betydelig høyere andeler av voksne mannlige homofile rapporterte å bli oppmuntret eller tvunget til å ha samleie opp til 19 år (Cunningham 1994); Sammenlignet med kontrollgruppen ble det observert høyere frekvenser av homoseksuell preferanse hos unge menn som fikk seksuelle overgrep i barndommen (Johnson 1987; Finkelhor xnumx, 1984; Wyre inn Tate xnumx; Cunningham xnumx; Glasser 2001; Rind xnumx; Garcia xnumx; Arreola 2005; Beitchman xnumx; Jinich xnumx; Laumann xnumx; Lenderking 1997; Paul 2001; Tomeo 2001; Freund xnumx). Det kan konkluderes med at homoseksuell interesse, uavhengig av alderen på tiltrekningsobjektet, har en vanlig årsak. Blanchards studier har vist at SBE også sees blant homofile og bifile pedofile, det vil si at slike individer har eldre brødre (Bogaert 1997).
Lee et al. (2002) forsøkte å fastslå hvilke av flere risikofaktorer - emosjonelle overgrep mot barn, atferdsproblemer og seksuelle overgrep mot barn - som var assosiert med følgende: pedofili, ekshibisjonisme, seksuelt misbruk. Seksuelle overgrep mot barn var en spesifikk risikofaktor for pedofili. Andre relaterte faktorer (emosjonelle overgrep og atferdsproblemer) var ikke så nært forbundet med pedofili. I tillegg, gitt den klare sammenhengen mellom tilstedeværelsen av flere homoseksuelle søsken i familien og incest, bør incest vurderes som et mulig alternativ til biologiske forklaringer. Når en bror (vanligvis den eldre) viser homoseksuelle tendenser, risikerer andre brødre å bli forført eller voldtatt, noe som kan fikse deres homoseksuelle aktivitet (Cameron 1995). I følge britisk statistikk forekommer 38% av tilfellene av seksuell vold i familien fra brorens side (Cawson xnumx). I følge forskeren Bartlett (2018), diskusjoner i populærpsykologi om hvorvidt en voksnes personlighet dannes avhengig av rekkefølgen på hans fødsel, er en lang historie med en stor mengde vitenskapelig litteratur som dekker tusenvis av publiserte verk (Damian xnumxa; Paulhus 2008; Laks xnumx). I løpet av de siste tiårene har forskning på dette problemet bygd på forestillingen om at konkurranse mellom brødre og søstre om ressursen til foreldrenes oppmerksomhet fører til det faktum at fødselsrekkefølgen til barn i familien påvirker barns individuelle egenskaper. Siden barn tilpasser seg bruken av forskjellige nisjer i familien, som regel, er eldre barn mer dominerende og tar over en del av foreldrenes krefter, mens senere barn er mer utadvendte og omgjengelige (Sulloway 1996). Det skal bemerkes at siden den varierende familiestørrelsen og sosioøkonomisk status i kombinasjon med små prøver i betydelig grad påvirker resultatene fra statistiske beregninger, bør studier der det er mulig å mer eller mindre tilstrekkelig studere ESBs sammenligning, inneholde minst 30 tusen søsken sammenligninger, mens hvordan studier som sammenligner relativt ensartede prøver fra familier anses som tilstrekkelige med utgangspunkt i 500-familier (Paulhus 2008). Selv om studier med små prøver viser motstridende data om ESB, i store studier (f.eks. Rohrer xnumx, n = 20 000; Damian xnumxb, n = 377 000), fødselsordningens påvirkning på individuelle kvaliteter (Damian xnumxa). Det disse empiriske dataene viser er en godt reproduserbar effekt der intelligensindikatorene til hvert påfølgende barn faller med omtrent en tidel av standardavviket hvis barnet lever til voksen alder (Kristensen 2007), som tydelig viser at årsaken til effekten er en reduksjon i foreldreinvestering, og ikke biologiske intrauterine prosesser. Storskala studier avdekker også effekten av fødselsrekkefølge på kvaliteter som akademisk ytelse, økonomisk suksess og selvmordsrisiko (Bjørngaard 2013; Svart xnumx).
Dermed har ikke det biologiske grunnlaget for attraksjon av samme kjønn, fremmet av hypotesen om brødrenes fødselsrekkefølge, noen empirisk støtte, mens det er mye empirisk bevis mot det.
Duality of LGBT + Attitude - Blanchard Movement
Anta at ESB og mødreimmunisering skjer og forårsaker endringer i atferd. I dette tilfellet kombinerer Blanchards hypotese homofili og transseksualisme (så vel som homofil pedofili) - og i den moderne "LHBT +" -bevegelsen er dette blasfemi. Ifølge American Psychological Association er for eksempel seksuell lyst og seksuell identitet helt ubeslektede fenomener (APA 2011 / 2014). I følge Blanchards hypotese er transseksualisme en patologi som er forårsaket av (1) en ekstrem manifestasjon av homoseksuell tiltrekning, der "feminiseringen" av hjernen er så uttalt at den også påvirker seksuell selvidentifisering; eller (2) et mentalt avvik der seksuell tiltrekning ikke er rettet mot det motsatte kjønn, men mot seg selv i bildet av det motsatte kjønn (Blanchard kalte den siste tilstanden "autogynephilia"14) (Blanchard 1989; Bailey 2003). Blanchard anser utvetydig transseksualisme som et patologisk fenomen. I et intervju bemerket Blanchard dessuten:
“... Jeg vil si at hvis det var mulig å starte helt fra bunnen av og ignorere hele historien om utelukkelse av homoseksualitet fra DSM, er normal seksualitet alt som er assosiert med reproduksjon.15... "(Cameron 2013).
En slik dristig posisjon medfører misnøye blant representantene for “LHBT +” - bevegelsen, spesielt i den delen av den som representerer “T” (Wyndzen xnumx; Troadsmap; Dreger 2008; Serano 2010).
Aktivister av “LGBT +” skriver om Blanchard - bevegelser:
“… Blanchard siteres ofte av anti-LHBT-grupper (…) Og hvorfor ikke? Blanchard vokste opp som katolikk, han har et veldig tradisjonelt syn på at ethvert samleie som ikke involverer penis og skjede er unormalt (...) Hvis Dr. Blanchard var nøtteaktig uten posisjon og autoritet, kunne han lett bli miskreditt. Men dette er ikke tilfelle - tvert imot, han var i JSM-komiteen med ansvar for parafilier og seksuelle dysfunksjoner (...) Han motarbeider åpent HBT-personer ... "(Tannehill xnumx).
På den annen side, bekreftelse av Blanchards hypotese stiller tvil om en av de grunnleggende dogmer av “LHBT +” - bevegelse - begrepet normativitet om mangfoldet av seksuell tiltrekning etter kjønn av et objekt. I dette tilfellet vil grunnen til homoseksuell tiltrekning bli avslørt - patologisk immunrespons. Ellers vil aktivister av "LGBT +" -bevegelsen måtte forvrenge forståelsen av medisin og biologi på en slik måte at de beregner immunresponsen som forårsaker spontanabort, vekttap, reduserte reproduksjonssjanser, en endring i den psyko-intellektuelle tilstanden som krever hormonelle medisiner og kirurgisk inngrep, samt pedofile preferanser og en tendens til vold er normalternativet.
I tillegg vil det være utsikter til forebygging av homoseksuelle preferanser hos gutter i analogi med bruk av anti-Rhesus immunoglobuliner i Rh-konflikt graviditet. Hvilken del av fremtidige foreldre, selv de som er lojale mot "LHBT +" -bevegelsen, vil bevisst nekte muligheten til å redusere risikoen for homofil attraksjon hos guttene? I dagens tid blir hver kvinne faktisk nøye forklart om abortens tillatelighet og rutine. Vil en kvinners rett til å påvirke fosterets liv også omfatte retten til å påvirke sin fremtidige seksuelle oppførsel, eller vil det være valgforbud og rettsforfølgelse av de profesjonelle som vil gi en slik mulighet?
En eller annen måte for øyeblikket er disse problemene sannsynlige.
Tolkningsproblemer
Det er noen viktige interne begrensninger i resultatene fra empiriske studier, likt de som ble diskutert i tidligere seksjoner. Å ignorere disse begrensningene er en av hovedårsakene til feiltolkning av forskning i det offentlige rom. Det er ganske fristende å anta, som vist med eksemplet med strukturen i hjernen, at hvis en bestemt biologisk profil er assosiert med en eller annen atferdsmessig eller psykologisk egenskap, så er en slik biologisk profil årsaken til denne egenskapen. Denne begrunnelsen er basert på feil.
Vi illustrerer kort noen av begrensningene som ligger i dette forskningsområdet ved å bruke følgende hypotetiske eksempel. Anta at vi må gjennomføre en sammenlignende studie av hjernen til yogainstruktører og kroppsbyggere. Hvis du søker lenge nok, vil det til slutt være statistisk signifikante forskjeller i ethvert område av den morfologiske strukturen eller hjernefunksjonene mellom disse gruppene. Men dette vil ikke bety at slike forskjeller bestemmer egenskapene til livsbanene til en yogainstruktør og en kroppsbygger. Hjerneegenskaper kan være resultatet snarere enn årsaken til særegne atferdsmønstre og interesser. Undersøkelser av nevroplastisitet viser at til tross for tilstedeværelsen av kritiske utviklingsperioder der hjernen endrer seg raskere og sterkere (for eksempel under språklig utvikling hos små barn), fortsetter hjernen å endre seg gjennom livet, og reagerer på atferdsmønstre (for eksempel å sjonglere eller leke på musikkinstrument), livserfaring, psykoterapi, medikamenter, psykologiske traumer og forhold. For Doidge 2007 for en nyttig og tilgjengelig oversikt over nevroplastisitetsstudier.
Å avgjøre om noe har en biologisk grunn er en ekstremt kompleks prosess, og å identifisere en spesifikk genetisk kobling er en enda vanskeligere oppgave. Studier som deklarativt gir ubestridelig ”bevis” på at homoseksuelle er ”født på den måten” er i beste fall inkonsekvente, og resultatene er i stor grad korrelert i sin natur.
I noen tilfeller, for eksempel i tvillingstudier, tyder bevis på at tidlige miljøfaktorer har en dominerende innflytelse på forekomsten av homoseksuelle tendenser. Korrelasjonen mellom de to faktorene betyr overhode ikke at det er en årsakssammenheng mellom dem. Basketballspillere er høye - spiller basketball korrelerer absolutt med høy vekst. Det er imidlertid ikke noe "basketballgen". Det er klart, noen interessante korrelasjoner blir presentert som påståtte årsaksfaktorer for politiske og propagandaformål.
Til slutt, antar at noen mennesker kan være disponert for homoseksuelle tendenser på grunn av genetisk, prenatal, hormonell påvirkning, eller andre fysiske eller hjerneegenskaper. Betyr dette at homofili er et medfødt fenomen? Slett ikke i forståelsen av hvordan dette er representert av media og populærkultur. Sjenke og kunstneriske unge gutter hvis far ikke ga oppmerksomhet til oppdragelse, ikke var et eksempel på riktig maskulin type oppførsel, kan risikere å utvikle homoseksuelle tilbøyeligheter. Dette skyldes ikke det homoseksuelle "genet", men på grunn av den forstyrrede mentale prosessen med dannelse av seksuell identitet. Slike gutter har et emosjonelt behov for selvbekreftelse og mannlig oppmerksomhet. Et lignende bilde er observert hos jenter som ikke samsvarer med klassiske seksuelle profiler. Problemene og emosjonelle behovene til slike barn spilles ofte av nåværende trender i det seksuelle og seksuelle verdensbildet.
Disse eksemplene illustrerer et av de vanlige problemene som oppstår med den utbredte tolkningen av slike studier - antakelsen om at nevrobiologiske faktorer bestemmer en spesifikk atferdsmodell.
Hvis naturen gir noen attraksjoner av samme kjønn, så hvorfor gir den ikke fysiske egenskaper som er nødvendige for å realisere den? For eksempel en tett og flerlags epitelmembran i endetarmen, i stand til å motstå langvarig friksjon, med kjertler som frigjør rikelig smøring, en tynnere penis for penetrering i endetarmen, etc. Hvis disse kjennetegnene var til stede blant homoseksuelle, kunne man snakke om medfødthet. Hvis de har et normalt sett med kromosomer og et normalt forplantningssystem, blir tiltrukket av et objekt som det ikke er mulig å bruke det til deres tiltenkte formål, virker foredraget om den biologiske tilstanden til dette fenomenet veldig spekulativt.
Meningen fra noen representanter for "LHBT +" -bevegelsen
American Psychological Association i 2014 ga ut en guide til psykologiske sykdommer og sexologi. Her er direkte sitater fra det:
"... For øyeblikket er ingen gener identifisert som kan være assosiert med homofili ..." (Rosario i APA 2014, s. 579)
"... Den ubestridelige virkeligheten er at menneskelig seksuell atferd bestemmes av en kombinasjon av mange faktorer: biologisk, sosial og valgfaktoren ..." (Kleinplatz i APA 2014, s. 256).

Forfatteren av flere kapitler fra APA-ledelsen er medlem av APAs ekspertkomité, professor Lisa Diamond, som ikke legger skjul på sine homoseksuelle preferanser. Diamond er imot teorien om homofiliens genetiske betingelse. Hun er sikker på at oppgaven ”homoseksuelle ble født på den måten og ikke kan endre seg” er feil. I 2013-året uttalte Diamond i et foredrag på Cornell University:
"... Jeg mener at det skeive samfunnet burde slutte å si" vi ble født på denne måten og vi kan ikke endre ", og bruke dette slagordet i vår kamp ... Jeg tror at vi ikke lenger trenger dette argumentet og til og med gjør vondt, for i dag er det samlet et overbevisende volum. vitenskapelige data kjent til “den andre siden” så vel som oss ... ”(Diamond 2013).

Forfatteren til mange bøker om kunst og filosofi, som ikke legger skjul på sine preferanser av samme kjønn, amerikanske Camilla Paglia, slår fast:
“... Homofili er ikke normen. Tvert imot, det er en utfordring mot normen ... skeive teoretikere - denne skrumpne gjengen svindlere av frilastere - prøvde å gå en poststrukturalistisk kurs og hevdet at det ikke er noen norm, siden alt er tilfeldig og relativt. Dette er den dumme blindveien der folk besatt av ord faller når de er døve, dumme og blinde for verden rundt dem. Naturen eksisterer, enten forskere liker det eller ikke, men i natur er forplantning den eneste og uforsonlige regelen. Dette er normen. Kjønnenes kropper er laget for reproduksjon. Penis passer til skjeden, og ingen bisarr sjonglering av ord kan endre dette biologiske faktum ... Ingen er født homoseksuelle. Ideen i seg selv er latterlig ... Homofili er en tilpasning, ikke en medfødt eiendom ... "(Paglia 1994, sider 70 - 76).
En annen fremtredende amerikansk aktivist, Cynthia Nixon, ble angrepet av LGBT +, en bevegelse for åpent å uttrykke synet om at hennes samme kjønnsdriv er drevet av personlig valg, ikke biologi (Witchell 2012).
LHBT + amerikansk aktivist - bevegelsesjournalist Brandon Ambrosino uttalte også at han ikke var født, men bevisst valgte en homoseksuell livsstil (Ambrosino 2014), noe som provoserte indignasjonen fra noen av kollegene i "LGBT +" -bevegelsen (Arana xnumx).

Kilde: Frazer Harrison / WireImage
Feminist og LHBT + aktivist - Karl Mantilla Movement skriver i sin artikkel:
“... Jeg har lenge trodd at LGBT + -strategien - bevegelsen for å bruke det medfødte argumentet er utrolig halt ... Selvfølgelig er dette et valg - hvordan kan det være ellers? … For en stund deltok jeg i en støttegruppe for kvinner som bestemte seg for å bli lesbiske i et tradisjonelt ekteskap. På et tidspunkt stilte jeg spørsmålet: "Hvordan forsto du at du er lesbisk?" En kvinne svarte at hun aldri følte seg følelsesmessig nær menn og at hun alltid ble bedre forstått av kvinner. En annen sa umiddelbart at også hun følte at hun bare kunne være følelsesmessig åpen med kvinner. De andre nikket enig. Hva var galt i den situasjonen? Nesten alle kvinner har det slik! Hver heterofil kvinne jeg noen gang har kjent, følte meg mer komfortabel med å stole på vennene sine, følte seg nærmere dem, følte seg bedre forstått og mer åpen for kvinner. Hvis det er det som trengs for å være lesbisk, så er alle kvinner lesbiske. Dette er like gammelt som verden ... kvinnens klager over at mennene deres ikke snakker med dem, ikke forstår deres følelser og ikke er interessert i det de sier. Noen av de vanligste artiklene i kvinneblader er hvordan du får mannen din til å åpne seg og snakke med deg ... følelsen av følelsesmessig nærhet til en person har ikke noe biologisk grunnlag, det er på grunn av en persons emosjonelle og psykologiske egenskaper ... over tid ble det klart for meg at kvinner i denne støttegruppen følte rett og slett enorm skyld for å ha forlatt sine ektemenn ... Så ideen om at de ikke kunne gjøre noe med det faktum at de var lesbiske, at det var en biologisk grunn, frigjorde dem fra skyld og ansvar for deres handlinger ... "(Mantilla xnumx).
LGBT + -aktivist, en California-basert bevegelse ved navn Gail Madwin, har til og med opprettet et helt nettsted som argumenterer for at homoseksuell atferd ikke er medfødt, men på grunn av et bevisst valg (Queer by choice). Tidligere LGBT + -aktivist David Benkof underbygger også det faktum at en homoseksuell livsstil på ingen måte bestemmes av noen biologiske faktorer (Benkof xnumx).
Merknader
1: vi ble født på den måten
2 Generelt ikke relatert til hverandre
3 Etter det "strenge" kriteriet om homoseksuelle tilbøyeligheter: 2 og mer av den såkalte Kinsey skala.
4 engelsk GWAS, Genome-Wide Association Studies
5 i det vitenskapelige samfunnet vedtok praksisen med å sende inn CV på konferanser - en kort artikkel, vanligvis 150 - 250 ord i størrelse - etterfulgt av publisering av en full artikkel i et tidsskrift
6 Engelsk: sannsynligvis født med en predisposisjon
7 I denne forbindelse kan fordelingen av resultater per person være begrenset
8 virilisering - en medisinsk betegnelse for et brudd der kvinnelige seksuelle egenskaper utvikler seg til mannlige
9 engelsk: “interstitielle kjerner i den fremre hypothalamus (INAH)”
10 English: “prepulse inhibition of the human startle response (PPI)”
11 Engelsk: “Faternal fødselsordreffekt (FBO)”
12 Se seksjonen om tvillingforskning
13 I tillegg er antigener i tilfelle av PK og transplantasjonsavstøtningsreaksjoner individuelle (faderlige i tilfelle av PK), men karakteristiske for hannen.
14 fra gresk biler - “selv-”, gini - “kvinne” og filia - “kjærlighet”; "Kjærlighet for seg selv som kvinne"
15 Jeg vil si at hvis man kunne starte fra bunnen av, ignorere all historien med å fjerne homoseksualitet fra DSM, er normal seksualitet det som er relatert til reproduksjon.
mer informasjon
Mer informasjon og detaljer finner du i følgende kilder:
1. Whitehead NE, Whitehead BK. Genene mine fikk meg til å gjøre det! Homoseksualitet og vitenskapelige bevis. Whitehead Associates. Utgave 5th 2018.
2. Mayer LS, McHugh PR. Seksualitet og kjønn: Funn fra biologiske, psykologiske og samfunnsvitenskapelige. The New Atlantis, Antall 50, Høst 2016.
3. Sprigg P., et al. Å få det rett ut: hva forskningen viser om homofili. Washington: Family Research Council (2004).
3. Harrub B, Thompson B, Miller D. “Dette er slik Gud skapte meg”, en vitenskapelig undersøkelse av homoseksualitet og “homofil gen”. Fornuft og åpenbaring. August 2004; 24 (8): 73.
5. Sorba r. ”Born Gay” Hoax. Ryan Sorba Inc. Første utgave 2007.
6. Whitehead NE. Et antistoffantistoff? Re-undersøkelse av immunhypotesen til mors. Journal of Biosocial Science 2007.
7. Ridder r. Født eller avlet? Vitenskap støtter ikke påstanden om at homofili er genetisk... Kultur & familieinstitutt. Bekymrede kvinner for Amerika. 2004.
8. van den Aardweg G. Homofili og biologiske faktorer: Ekte bevis - Ingen; Villedende tolkninger: Rikelig. Reprinted fra NARTH Bulletin, Winter 2005.
9. Hubbard R, Wald E. Eksplodere genmyten: Hvordan genetisk informasjon produseres og manipuleres av forskere, leger, arbeidsgivere, forsikringsselskaper, lærere og rettshåndhevere. Beacon Press, Boston; 1999.
Bibliografiske kilder
- Vasilchenko G.S. Sexopatologi: Håndbok / Ed. GS Vasil. - M., 1990.
- Yarygin V.N. (2003) // Biologi. I boka 2 Ed. VN Yarygin / Yarygin V.N., Vasilieva V.I., Volkov I.N., Sinelshchikova V.V. 5 utg., Rev. og legg til. - M .: Higher School, 2003. Book 1 - 432s., Book 2 - 334s.
- ASHG 2015. Epigenetisk algoritme forutsier nøyaktig mannlige seksuelle orienteringsfunn rapportert på ASHG 2015 årsmøte. For øyeblikkelig utgivelse torsdag oktober 8, 2015 http://www.ashg.org/press/201510-sexual-orientation.html
- Albrecht ED, Pepe GJ. Østrogenregulering av placentaangiogenese og fosterets ovarieutvikling under primatgraviditet, ”International Journal of Developmental Biology 54, nr. 2 - 3 (2010): 397 - 408, http://dx.doi.org/10.1387/ijdb.082758ea
- Allen S. Den problematiske jakten på en 'homofil gen'. The Daily Beast. 20.11.2014. https://www.thedailybeast.com/the-problematic-hunt-for-a-gay-gene (01.12.2017 bekreftet)
- Ambrosino B. Jeg ble ikke født på denne måten. Jeg velger å være homofil. Den nye republikken. 28. januar 2014. https://newrepublic.com/article/116378/macklemores-same-love-sends-wrong-message-about-being-gay
- APA. Amerikansk psykologforening. Svar på spørsmålene dine. Om transpersoner, kjønnsuttrykk og kjønnsidentitet. Utarbeidet av kontoret for PR og medlemmer av foreningen. Trykt 2011; oppdatert 04 / 2014.https: //www.apa.org/topics/lgbt/transgender-russian.pdf
- Arana G. Ezra Kleins Queer New Hire. 13. mars 2014. The American Prospect.
- Arreola, SG, Neilands, TB, Pollack, LM, Paul, JP & Catania, JA (2005) Høyere forekomst av seksuelle overgrep i barndommen blant latino menn som har sex med menn enn ikke-latino menn som har sex med menn: data fra Urban Men's Health Study. Barnemishandling og forsømmelse 29, 285-290.
- Bailey J. M, et al., "A Test of the Mother's Stress Theory of Human Male Homosexuality," Archives of Sexual Behaviour 20, nr. 3 (1991): 277 - 293, http://dx.doi.org/10.1007/BF01541847
- Bailey, J. Michael (2003). Mannen som ville bli dronning: Vitenskapen om kjønnsbending og transseksualisme. Joseph Henry Press
- Bailey JM, et al. Genetiske og miljømessige påvirkninger på seksuell legning og dens korrelasjoner i en australsk tvillingprøve. J Pers Soc Psychol. 2000 Mar; 78 (3): 524-36.
- Bains JS, Wamsteeker Cusulin JI, Inoue W. Stressrelatert synaptisk plastisitet i hypothalamus. Nat Rev Neurosci. 2015 Jul; 16 (7): 377-88. doi: http://dx.doi.org/10.1038/nrn3881
- Baron M. Genetikk og seksuell legning. Biologisk psykiatri. Juni 1 - 15, 1993, Volume 33, Issues 11-12, Pages 759 - 761.
- Bartlett NT, Hurd PL.Faderlig fødselspåvirkning på personlighet: Vil rimelige krav kreve ekstraordinært bevis? Arch Sex Behav. 2018 Jan; 47 (1): 21-25. doi: 10.1007 / s10508-017-1109-z.
- Be, G., Velasquez, P. & Youlton, R. (1997) Spontan abort: cytogenetisk studie av 609 tilfeller. Revista Medica de Chile 125, 317-322.
- Bearman PS, Brückner H. Motsatt - Sex tvillinger og ungdommer samme - sexattraksjon. American Journal of Sociology 2002 107: 5, 1179-1205
- Bearman, PS, & Brückner, H. (2002). Motsatt kjønn tvillinger og ungdoms samme kjønn attraksjon. American Journal of Sociology, 107, 1179-1205. doi: 10.1086 / 341906.
- Beer, AE & Billingham, RE (1975) Immunologiske fordeler og farer med melk i mors-perinatal forhold. Arkiv for indremedisin 83, 865-871.
- Beitchman, JH, Zucker, KJ, Hood, JE, Da Costa, GA & Akman, S. (1991) En gjennomgang av kortsiktige effekter av seksuelt misbruk av barn. Barnemishandling og forsømmelse 15, 537–556.
- Benkof D. Ingen er "født på den måten," sier homofile historikere. The Daily Caller. 19.03.2014. dailycaller.com/2014/03/19/nobody-is-born-that-way-gay-historians-say/
- Ben-Porath, Y., et al. (1976) Har egentlig sexpreferanser noe? QJ Econ. 90, 285 - 307.
- Berenbaum SA. Hvordan hormoner påvirker atferds- og nevral utvikling: Introduksjon til den spesielle saken om 'Gonadal hormoner og kjønnsforskjeller i atferd. Developmental Neuropsychology 14 (1998): 175 - 196, http://dx.doi.org/10.1080/87565649809540708
- Biggar, RJ, et al. (1999) Kjønnstall, familiestørrelse og fødselsrekkefølge. Am. J. Epidemiol. 150, 957 - 962.
- Billings, Beckwith. Technology Review. Juli 1993, s. 60.
- Bjørngaard, JH, Bjerkeset, O., Vatten, L., Janszky, I., Gunnell, D., & Romundstad, P. (2013). Mødrealder ved fødsel av barn, fødselsordre og selvmord i ung alder: A søsken sammenligning. American Journal of Epidemiology, 177, 638-644. https://doi.org/10.1093/aje/kwt014.
- Black, SE, Devereux, PJ, & Salvanes, KG (2005) Jo mer jo bedre? Effekten av familiestørrelse og fødselsrekkefølge på barns utdanning. Quarterly Journal of Economics, 120, 669-700. https://doi.org/10.2307/25 098749.
- Blanchard R (august 1989). "Klassifiseringen og merkingen av ikke-homoseksuelle kjønnsdysforier." Arkiv for seksuell atferd. 18 (4): 315–34. doi:10.1007/BF01541951
- Blanchard R, Bogaert AF. (1996) Homoseksualitet hos menn og antall eldre brødre. American Journal of Psychiatry 153, 27 - 31.
- Blanchard R, Bogaert AF. Homoseksualitet hos menn og antall eldre brødre. The American Journal of Psychiatry; Jan 1996a; 153, 1; Research Library, s. 27
- Blanchard R., et al. (2000) Fostersk fødselsrekkefølge og seksuell legning hos pedofile. Buer seksuell atferd 29, 463 - 478.
- Blanchard, R. & Bogaert, AF (1996b) Biodemografisk sammenligning av homofile og heterofile menn i Kinsey Interview-dataene. Arkiv for seksuell atferd 25, 551-579.
- Blanchard, R. & Bogaert, AF (1998) Fødselsrekkefølge hos homofile versus heterofile seksualforbrytere mot barn, pubescents og voksne. Arkiv for seksuell atferd 27, 595-603.
- Blanchard, R. & Ellis, L. (2001) Fødselsvekt, seksuell orientering og kjønnet til foregående søsken. J. biosoc. Sci. 33, 451-467.
- Blanchard, R. (2014). Oppdage og korrigere for forskjeller i familiestørrelse i studien av seksuell legning og fødselsrekkefølge fra brødrene Archives of Sexual Behaviour, 43, 845 - 852. https://doi.org/10.1007/s10508-013-0245- 3.
- Blanchard, R. Fraternal fødselsrekkefølge, familiestørrelse og mannlig homoseksualitet: Metaanalyse av studier som spenner over 25 år. Arch Sex Behav (2018) 47: 1. https://doi.org/10.1007/s10508-017-1007-4
- Blanchard, R., & Bogaert, AF (2004). Andel homofile menn som skylder sin seksuelle orientering til broderlig fødselsrekkefølge: Anestimert basert på uavhengige sannsynlighetsprøver. American Journal of Human Biology, 16, 151-157.
- Blanchard, R., & VanderLaan, DP (2015). Kommentar til Kishida og Rahman (2015), inkludert en metaanalyse av relevante studier om broderlig fødselsrekkefølge og seksuell legning hos menn. Archives of SexualBehavior, 44, 1503-1509. doi: 10.1007 / s10508-015-0555-8
- Blanchard, R., Barbaree, HE, Bogaert, AF, Dickey, R., Klassen, P., Kuban, ME & Zucker, KJ (2000) Broderlig fødselsrekkefølge og seksuell orientering hos pedofile. Arkiv for seksuell atferd 29, 463–478.
- Blokk N, "Hvordan arvelighet misviser om rase," Erkjennelse 56, nr. 2 (1995): 103 - 104, http://dx.doi.org/10.1016/0010-0277(95)00678-R
- Bogaert, AF (2003). Interaksjon mellom eldre brødre og sex-typing i prediksjonen om seksuell legning hos menn. Arkiv for seksuell atferd, 32, 129 - 134.
- Bogaert, AF (2004). Utbredelsen av mannlig homoseksualitet: Effekten av broderlig fødselsrekkefølge og variasjoner i familiens størrelse. Journal of Theoretical Biology, 230, 33 - 37.
- Bogaert, AF (2005). Kjønnsrolle / identitet og søskenforhold hos homofile. JournalofSexandMaritalTherapy, 31,217 - 227. https: // doi. org / 10.1080 / 00926230590513438.
- Bogaert, AF (2006) Biologiske versus ikke-biologiske eldre brødre og mannlig seksuell legning. Proceedings of the National Academy of Sciences 103, 10771 - 10774.
- Bogaert, AF, Bezeau, S., Kuban, M. & Blanchard, R. (1997) Pedofili, seksuell orientering og fødselsrekkefølge. Journal of Abnormal Psychology 106, 331-335.
- Bogaert, AF, & Skorska, M. (2011). Seksuell orientering, broderens fødselsrekkefølge og mors immunhypotese: Areview. Frontiers in Neuroendocrinology, 32, 247-254.
- Bogaert, AF (2005). Søkeforhold mellom kjønn og seksuell legning hos menn og kvinner: Nye tester i to nasjonale sannsynlighetsprøver. Archives of Sexual Behaviour, 34, 111 - 116. doi: 10.1007 / s10508-005-1005-9.
- Bogaert, AF (2010). Fysisk utvikling og seksuell orientering menn og kvinner: En analyse av NATSAL-2000. Arkiv for seksuell atferd, 39, 110 - 116.doi: 10.1007 / s10508-008-9398-x.
- Briggs WM. På antatt nyoppdagede “homofile gener.” Eller betydningen av modellferdighet. Oktober 13, 2015. wmbriggs.com/post/17053/
- Byne W, Tobet S, Mattiace LA, et al. De interstitielle kjernene til den menneskelige fremre hypothalamus: en undersøkelse av variasjon med sex, seksuell legning og HIV-status. Horm Behav. 2001 Sep; 40 (2): 86-92. http://dx.doi.org/10.1006/hbeh.2001.1680
- Byne W. Det biologiske beviset utfordret. Scientifiс American, Mai 1994, s. 50 - 55.
- Caldwell, JC (1997). Nå en stabil global befolkning: Hva vi har lært, og hva vi må gjøre. Health Transition Review, 7, 37 - 42.
- Cameron P, et al. Forårsaker incest homoseksualitet? Psykologiske rapporter, 1995, 76, 611-621.
- Cameron L. Hvordan psykiateren som skrev om bruksanvisningen om sexprat om sex? Kortet. Apr 11 2013. https://motherboard.vice.com/en_us/article/ypp93m/heres-how-the-guy-who-wrote-the-manual-on-sex-talks-about-sex
- Cantor, JM, Blanchard, R., Paterson, AD & Bogaert, AF (2002) Hvor mange homofile menn skylder sin broderlige fødselsordre sin seksuelle legning? Arkiv for seksuell atferd 31, 63–71.
- Cardwell, CR, Carson, DJ & Patterson, CC (2005) Foreldres alder ved fødsel, fødselsrekkefølge, fødselsvekt og svangerskapsalder er assosiert med risikoen for barndom Type 1-diabetes: en britisk regional retrospektiv kohortstudie. Diabetisk medisin 22-200.
- Cawson P, et al. Mishandling av barn i Storbritannia: en studie av forebygging av misbruk og omsorgssvikt. NSPCC forskningsresultater november 2000.
- Chaix, R., Cao, C., og Donnelly, P. (2008). Er valg av kompis hos mennesker MHC-avhengig? PLoS Genetics, 4, e1000184.
- Cohen-Kettenis PT, kjønnsendring i 46, XY personer med 5α-Reductase-2 Mangel og 17β-Hydroxysteroid Dehydrogenase-3 Mangel. Arkiv for seksuell atferd 34, nr. 4 (2005): 399 - 410, http://dx.doi.org/10.1007/s10508-005-4339-4
- Collins FS Guds språk. New York, NY Simon & Schuster, Inc. 2006.
- Cote, K., Earls, CM & Lalumiere, ML (2002) Fødselsrekkefølge, fødselsintervall og avvikende seksuelle preferanser blant sexforbrytere. Sexual Abuse: a Journal of Research and Treatment 14, 67–81.
- Cunningham, RN, et al. (1994) Forbindelsen av fysisk og seksuelt misbruk med HIV-risikoatferd i ungdomstiden og ung voksen alder: implikasjoner for folkehelsen. Overgrep mot barn Negl. 18, 233 - 245.
- Damian, RI, & Roberts, BW (2015a). Setter debatten om fødselsrekkefølge og personlighet. Proceedings of the National Academy of Sciences, 112, 14119-14120. https://doi.org/10.1073/pnas.1519064112.
- Damian, RI, & Roberts, BW (2015b). Foreningene av fødselsordre med personlighet og intelligens i et representativt utvalg av amerikanske videregående studenter. Journal ofResearchinPersonality, 58,96-105.https: // doi.org/10.1016/j.jrp .2015.05.005.
- Dankers, MK, Roelen, D., Korfage, N., de Lange, P., Witvliet, M., Sandkuiil, I., Doxiadis, II & Claas, FH (2003) Differensiell immunogenisitet av faderlige HLA klasse I-antigener hos gravide kvinner. Human Immunology 64, 600-606.
- Davis N. Naturlige fødte mordere: mennesker disponert for drap, antyder studien. The Guardian. 28.09.2016. https://www.theguardian.com/science/2016/sep/28/natural-born-killers-humans-predisposed-to-study-suggests (01.12.2017 Verified)
- Dawkins R. A Devil's Chaplain: Reflections on Hope, Lies, Science, and Love. First Mariner Books utgave 2004
- Diamond Lisa. Hvor forskjellig er seksuell orientering mellom kvinner og menn? 17.10.2013/2/43. Cornell University. https://www.youtube.com/watch?v=m13rTHDOuUBw&feature=youtu.be&t=01.12.2017mXNUMXs (Hentet XNUMX)
- Doidge Norman, hjernen som endrer seg: Historier om personlig triumf fra frontene til hjernevitenskap (New York: Penguin, 2007)
- Dörner Günter et al., "Stressful Events in Prenatal Life of Bi- og Homosexual Men," Eksperimentell og klinisk endokrinologi 81, nr. 1 (1983): 83 - 87, http://dx.doi.org/10.1055/s-0029-1210210
- Drabant EM et al., "Genome-Wide Association Study of Sexual Orientation in a Large, Web-based Cohort," 23andMe, Inc. (Programnummer: 2100W) Presentert på 62. årsmøte for The American Society of Human Genetics, 7. november 2012 i San Francisco, California. http://abstracts.ashg.org/cgi-bin/2012/ashg12s?author=drabant&sort=ptimes&sbutton=Detail&absno=120123120&sid=320078
- Dreger AD. Kontroversen som omgir mannen som ville bli dronning: En sakshistorie om politikk for vitenskap, identitet og sex i internettalderen. Arkiv for seksuell atferd. 2008; 37 (3): 366-421. doi: 10.1007 / s10508-007-9301-1.
- Ebstein Richard P. et al., “Genetics of Human Social Behaviour,” Neuron 65, nr. 6 (2010): 831– 844, http://dx.doi.org/10.1016/j.neuron.2010.02.020
- Eiben, B., Bahr-Porsch, S., Borgman, S., Gatz, G., Gellert, G. & Goebel, R. (1990) Cytogenetisk analyse av 750 spontane aborter med den direkte forberedende metoden til korion villi og dens implikasjoner for å studere genetiske årsaker til svangerskapsavfall. American Journal of Human Genetics 47, 656-663.
- Eiben, B., Borgman, S., Schubbe, I. & Hansman, I. (1987) En cytogen studieretning fra chorionic villi på 140 spontane aborter. Human Genetics 77, 137-141.
- Ellis L, Blanchard R (2001) Fødselsrekkefølge, forhold mellom søsken og kjønn og mødre hos mødre hos homoseksuelle og heterofile menn og kvinner. Personal. individ. Diffs 30, 543 - 552.
- Ellis Lee og Cole-Harding Shirley, "Effektene av prenatal stress og prenatal alkohol- og nikotineksponering, på menneskelig seksuell legning," Physiology & Behavior 74, nr. 1 (2001): 213-226, http://dx.doi.org/10.1016/S0031-9384(01)00564-9
- Ellis Lee et al., "Seksuell orientering av menneskelig avkom kan bli endret av alvorlig mors belastning under graviditet," Journal of Sex Research 25, nr. 2 (1988): 152 - 157, http://dx.doi.org/10.1080/00224498809551449
- Ennis D. Menneskerettighetskampanje setter syn på Johns Hopkins etter kontroversiell trans-rapport. 2016. NBC News.
- Evdokimova, VN, Nikitina, TV, Lebedev, IN, Sulchanova, NN & Nazarenko, SA (2000) Kjønnsforhold i tidlig embryonal dødelighet hos mennesker. Ontogenez 31, 251-257.
- Fausto-Sterling A., Balaban E. Genetikk og seksuell orientering av menn. Science. 1993; 261: 1257. http://dx.doi.org/10.1126/science.8362239
- Finkelhor, D. (1979) Seksuelt utsatte barn. Free Press, New York.
- Finkelhor, D. (1984) Seksuelt misbruk av barn: Ny teori og forskning. Free Press, New York.
- Finn R. Biologisk bestemmelse av seksuell oppvarming som et forskningsfelt. Forskeren 10 [1]: Jan. 08, 1996.
- Flannery, KA & Liderman, J. (1994) En test av den immunreaktive teorien for opprinnelsen til nevroutviklingsforstyrrelser hos avkom til kvinner med immunforstyrrelse. Cortex 30, 635-645
- Francis AM (2008). Familie- og seksuell legning: Familiedemografiske sammenhenger av homoseksualitet hos menn og kvinner.Journal of Sex Research, 45, 371 - 377. doi: 10.1080 / 00224490802398357.
- Freund, K. & Kuban, M. (1994) Grunnlaget for den misbrukte overgrepsteorien om pedofili: en videre utdyping av en tidligere studie. Arkiv for seksuell atferd 23, 553-563.
- Frisch, M., & Hviid, A. (2006) Barndomsfamilien korrelerer med heterofile og homofile ekteskap: En nasjonal kohortestudie av to millioner danskere. Archives of Sexual Behavior, 35,533-547. Doi: 10.1007 / s10508006-9062-2.
- Garcia, J., Adams, J., Friedman, L. & East, P. (2002) Koblinger mellom tidligere misbruk, selvmordstanker og seksuell orientering blant studenter i San Diego. Journal of American College Health 51, 9-14.
- Gasparoni, A., Avanzini, A., Ravagni Probizer, F., Chirico, G., Rondini, G. & Severi, F. (1992) IgG-underklasser sammenlignet i mors- og ledningsserum og morsmelk. Archives of Disease in Childhood 67 (1), spesialnummer, 41–43.
- Gavrilets S, Friberg U, Rice WR. Forstå homoseksualitet: gå videre fra mønstre til mekanismer. Arch Sex Behav. 2017. DOI 10.1007 / s10508-017-1092-4
- Gelman M. Gay-oppdatering av tabloid-hype. Statistisk modellering, årsakssammenheng og samfunnsvitenskap. Oktober 10, 2015. https://andrewgelman.com/2015/10/10/gay-gene-tabloid-hype-update/
- Ginalksi, K., Rychlewski, L., Baker, D. & Grishin, NV (2004) Proteinstrukturforutsigelse for den mannlige spesifikke regionen i det menneskelige Y-kromosomet. Proceedings of the National Academy of Sciences 101, 2305-2310
- Glasser, M., et al. (2001) Syklus av seksuelle overgrep mot barn: koblinger mellom å være et offer og å bli en gjerningsmann. Br. J. Psychiat. 179, 482 - 494.
- Gomes AR, Souteiro P, SIlva CG, et al. Utbredelse av testosteronmangel hos HIV-infiserte menn under antiretroviral terapi. BMC Infect Dis. 2016; 16: 628. Publisert online 2016 nov 3. http://dx.doi.org/10.1186/s12879-016-1892-5
- Greally J. Overtolket ukens studie av epigenetikk (2). EpgntxEinstein. Bloggen til Center for Epigenomics ved Albert Einstein College of Medicine i Bronx, New York City.
- Green, R. (2000) Fødselsrekkefølge og forhold mellom brødre og søstre i transseksuelle. Psykologisk medisin 30, 789 - 795.
- Gualteri, T. & Hicks, RE (1985) En immunreaktiv teori om selektiv mannlig lidelse. Oppfør deg. Brain Sci. 8, 427-477.
- Guleria I, Sayegh MH. Morslig aksept av fosteret: ekte menneskelig toleranse. J Immunol March 15, 2007, 178 (6) 3345-3351; DOI: https://doi.org/10.4049/jimmunol.178.6.3345
- Haler, A. & Fauzdar, A. (2006) Skjevt sexforhold og lav aneuploidi ved tilbakevendende tidlig savnet abort. Indian Journal of Medical Research 124, 9-10.
- Hall Lynn S. og Love Craig T., “Finger-Length Ratios in Female Monozygotic Twins Discordant for Sexual Orientation,” Archives of Sexual Behaviour 32, nr. 1 (2003): 23 - 28, http://dx.doi.org/10.1023/A:1021837211630
- Hamer D, Copeland P. The Desire of Desire: Search for the Gay Gen and the Biology of Behaviour. Simon og Schuster 1994
- Hamer D. The God Gen: How Faith is Hardwired into our Genes. Dobbeltdag xnumx
- Hamer DH et al., "En kobling mellom DNA-markører på X-kromosomet og mannlig seksuell legning," Science 261, nr. 5119 (1993): 321 - 327, http://dx.doi.org/10.1126/science.8332896
- Han, TH, Chey, MJ & Han, KS (2006) Granulocyttantistoffer i koreanske nyfødte med nøytropeni. Journal of the Korean Medical Society 21, 627-632.
- Harrison Halstead, "En teknisk kommentar til avisen, 'Seksuell orienteringsrelaterte forskjeller i forhåndshemming av menneskelig opprørt svar,'" University of Washington nettsted, 15 desember 2003, http://www.atmos.washington.edu/ ~ harrison / rapporter / rahman.pdf.
- Hatton GI. Funksjonsrelatert plastisitet i hypothalamus. Annu Rev Neurosci. 1997; 20: 375-97. http://dx.doi.org/10.1146/annurev.neuro.20.1.375
- Hoekzema E, et al. Graviditet fører til langvarige endringer i menneskets hjernestruktur. Nature Neuroscience volume 20, sider 287 - 296 (2017).
- Heston, L.L., Shields, J., "Homosexuality in Twins A Family Study and a Registry Study" Arch Gen Psychiat. 1968;18:149
- Hildebrand, H., Finkel, Y., Grahnquist, L., Lindholm, J., Ekbom, A. & Aksling, J. (2003) Endring av mønster for pediatrisk inflammatorisk tarmsykdom i Nord-Stockholm 1990–2001. Gut 52 1432– 1434.
- Hines M. Prenatal endokrine påvirkninger på seksuell orientering og på seksuelt differensiert barndomsatferd. Front Neuroendocrinol. 2011 Apr; 32 (2): 170 - 182. doi: 10.1016 / j.yfrne.2011.02.006
- Hines Melissa et al., "Prenatal stress og kjønnsrolleatferd hos jenter og gutter: En langsgående, befolkningsundersøkelse," Hormoner og atferd 42, nr. 2 (2002): 126 - 134, http://dx.doi.org/10.1006/hbeh.2002.1814
- Hönekopp J et al., “Forholdet mellom andre og fjerde siffer (2D: 4D) og kjønnshormonnivåer for voksne: Nye data og en metaanalytisk gjennomgang,” Psychoneuroendocrinology 32, nr. 4 (2007): 313 - 321, http://dx.doi.org/10.1016/j.psyneuen.2007.01.007
- Horgan, John. (1995) “Gay Genes, Revisited.” Scientific American, vol. 273, nei. 5, 1995, pp. 26 - 26. JSTOR, JSTOR, www.jstor.org/stable/24982058
- Hubbard R., Wald E. Eksploderende genmyten: Hvordan genetisk informasjon produseres og manipuleres av forskere, leger, arbeidsgivere, forsikringsselskaper, lærere og rettshåndhevere. 1999. Boston Press. ISBN: 978-080700431-9, på side 95 - 96.
- Huffpost 2017. Dean Hamer og Joe Wilson. https://www.huffingtonpost.com/author/qwavesjoe-855 (01.12.2017 bekreftet)
- Hughes IA, et al., "Androgen insensitivity syndrom," The Lancet 380, nr. 9851 (2012): 1419 - 1428, http://dx.doi.org/10.1016/S0140-6736%2812%2960071-3
- Human Genome Resources hos NCBI 2017. https://www.ncbi.nlm.nih.gov/projects/genome/guide/human/
- Izetbegovic S. Forekomst av ABO Og RhD Inkompatibilitet med Rh Negative Mothers. Materia Socio-Medica. 2013; 25 (4): 255-258. doi: 10.5455 / msm.2013.25.255-258.
- James WH. To hypoteser om årsakene til mannlig homofili og pedofili. J.biosoc.Sci, (2006) 38, 745 - 761, doi: 10.1017 / S0021932005027173
- James, WH (1975) Kjønnsforhold og kjønnsammensetning av eksisterende søsken. Ann. hum. Genet. 38, 371 - 378.
- James, WH (1985) Den påståtte forfølgende broreffekten i kjønnsforhold. Behav. Brain Sci. 8, 453.
- James, WH (1996) Bevis for at pattedyrens kjønnsforhold ved fødselen er delvis kontrollert av foreldrenes hormonnivå på unnfangelsestidspunktet. Journal of Theoretical Biology 180, 271 - 286.
- James, WH (2004) Årsaken (e) til den broderlige fødselseffekten hos mannlig homofili. Journal of Biosocial Science 36, 51 - 59, 61 - 62.
- James, WH (2004b) Ytterligere bevis på at pattedyrens kjønnsforhold ved fødselen er delvis kontrollert av foreldrehormonnivåer rundt unnfangelsestidspunktet. Menneskelig reproduksjon 19, 1250 - 1256.
- Jinich, S., Paul, JP, Stall, R., Acree, M., Kegeles, S., Hoff, C. & Coates, T. (1998) Barndoms seksuelle overgrep og HIV-risikovillig oppførsel blant homofile og bifile menn ... AIDS og atferd 2, 41-51.
- Johnson, RL & Shrier, DK (1987) Tidligere seksuell vold fra kvinner av mannlige pasienter i en ungdomsmedisinsk klinikkpopulasjon. Er. J. Psychiat. 144, 650-652.
- Juntunen, KS, Laara, EM & Kauppila, AJ (1997) Grand grand multiparity og fødselsvekt. Obstetrics and Gynecology 90, 495-499.
- Kallmann, Franz J., "Comparative Twin Study on the Genetic Aspects of Male Homosexuality," Journal of Nervous and Mental Disease 115, no. 4 (1952): 283 - 298
- Kano, T., Mori, T., Furudono, M., Kanda, T., Maeda, Y., Tsubokura, S., Ushiroyama, T. & Ueki, M. (2004) Kjønnsforskjeller mellom aborter og nyfødte hos kvinner med all-immun tilbakevendende aborter. Reproduktiv biomedisin online 9, 306-311.
- Kendler KS et al., "Seksuell orientering i en amerikansk nasjonal prøve av tvilling- og søskenpar i Twin," American Journal of Psychiatry 157, nr. 11 (2000): 1843 - 1846, http://dx.doi.org/10.1176/appi.ajp.157.11.1843
- Kishida, M., & Rahman, Q. (2015). Broderens fødselsrekkefølge og ekstrem rettferdighet som prediktorer for seksuell orientering og kjønnsavvik hos menn. Archives of Sexual Behavior, 44, 1493–1501. https: // doi. org / 10.1007 / s10508-014-0474-0.
- Kleinplatz & Diamond 2014, APA Handbook, Volume 1, s. 256-257
- Kolb B, Whishaw IQ. Hjernens plastisitet og atferd. Årlig gjennomgang av psykologi. Vol. 49: 43-64. https://doi.org/10.1146/annurev.psych.49.1.43
- Kranz F et al., "Ansiktsoppfatning moduleres av seksuell preferanse," Current Biology 16, nr. 1 (2006): 63 - 68, http://dx.doi.org/10.1016/j.cub.2005.10.070
- Kristensen, P., & Bjerkedal, T. (2007). Forklare forholdet mellom fødselsrekkefølge og intelligens. Vitenskap, 316, 1717. https://doi.org/10.1126/ science.1141493.
- Lalumiere, ML, Harris, GT, Quinsey, VL & Rice, ME (1998) Seksuell avvik og antall eldre brødre blant sexforbrytere. Seksuelt misbruk: en tidsskrift for forskning og behandling 10, 5-15.
- Långström Niklas et al., "Genetiske og miljømessige effekter på seksuell atferd av samme kjønn: En populasjonsstudie av tvillinger i Sverige," Archives of Sexual Behaviour 39, nr. 1 (2010): 75 - 80, http://dx.doi.org/10.1007/s10508-008- 9386-1.
- Lasco MS, et al., "Mangel på dimorfisme av sex eller seksuell orientering i den fremre menneskelige fremtid," Brain Research 936, nr. 1 (2002): 95 - 98, http://dx.doi.org/10.1016/S0006-8993(02)02590-8
- Laumann, EO, Gagnon, JH, Michaels, S. & Michael, RT (1993) Overvåking av AIDS og andre sjeldne populasjonshendelser: en nettverkstilnærming. Journal of Health and Social Behavior 34, 7-22.
- Lauterbach, MD, Raz, S. & Sander, CJ (2001) Hypoksisk risiko hos nyfødte hos premature fødte spedbarn: Påvirkningen av kjønn og alvorlighetsgraden av åndedrettsstress på kognitiv utvinning. Nevropsykologi 15, 411-420.
- Lee, JKP, et al. (2002) Utviklingsrisikofaktorer for seksuelle krenkelser. Overgrep mot barn Negl. 26, 73 - 92.
- Lee, RM & Silver, RM (2000) Gjentatt graviditetstap: oppsummering og kliniske anbefalinger. Seminarer i reproduktiv medisin 18, 433-440.
- Lenderking, WR, Wold, C., Mayer, KH, Goldstein, R., Losina, E. & Seage, GR (1997) Barnas seksuelle overgrep blant homofile menn. Prevalens og assosiasjon med usikker sex. Journal of General Internal Medicine 12, 250-253.
- Lenroot RK, Gogtay N, Greenstein DK, et al. Seksuell dimorfisme av hjerneutviklingsbaner under barndom og ungdom. NeuroImage. 2007; 36 (4): 1065-1073. doi: 10.1016 / j.neuroimage.2007.03.053.
- LeVay Simon, "En forskjell i hypotalamisk struktur mellom heterofile og homoseksuelle menn," Science 253, nr. 5023 (1991): 1034 - 1037, http://dx.doi.org/10.1126/science.1887219
- LeVay, S. (2016). Bra, rett og grunnen til: Vitenskapen om seksuell orientering (2nd utg.). Oxford, Storbritannia: Oxford University Press.
- Lippa Richard A., “Er 2D: 4D finger-lengde-forhold relatert til seksuell orientering? Ja for menn, nei for kvinner, ”Journal of Personality and Social Psychology 85, no. 1 (2003): 179 - 188, http://dx.doi.org/10.1037/0022-3514.85.1.179
- Lombardi, CM, & Hurlbert, SH (2009). Misbeskrivelse og misbruk av ensidige tester. Austral Ecology, 34, 447-468.
- Lykken, D.T., McGue, M., Tellegen, A., "Recruitment Bias in Twin Research: The Rule of TwoThirds Reconsidered" Behav. Genet. 1987;17:343
- MacCulloch, SI, Gray, NS, Phillips, HK, Taylor, J. & MacCulloch, MJ (2004) Fødselsrekkefølge hos sexkrenkende og aggressivt krenkende menn. Arkiv for seksuell atferd 33, 467–474.
- Magnus, P., Berg, K. & Bjerkedel, T. (1985) Foreningen av paritet og fødselsvekt: testing av sensibiliseringshypotesen. Tidlig menneskelig utvikling 12, 49–54
- Maguire EA, Gadian DG, Johnsrude IS, et al. Navigasjonsrelatert strukturell endring i hippocampi til drosjesjåfører. Fortsettelser av National Academy of Sciences of the United States of America. 2000; 97 (8): 4398-4403.
- Mainardi M, et al. Miljø, leptinfølsomhet og hypotalamisk plastisitet. Nevral plastisitet. 2013. Volum 2013 (2013), artikkel-ID 438072, 8 sider http://dx.doi.org/10.1155/2013/438072
- Manikkam, M., Crespi, EJ, Doop, DD, Herkimer, C., Lee, JS, Yu, S., Brown, MB, Foster, DL & Padmanabhan, V. (2004) Føtal programmering: prenatal testosteronoverskudd fører til forsinket fostervekst og postnatal oppvekst hos sau. Endokrinologi 145-790.
- Manning JT. (2001) Sifferforhold: en peker til fruktbarhet, atferd og helse. Rutgers University Press, London.
- Mantilla K. Biologi, rumpa mi. Av ryggen vår: en kvinners nyhetsjournal 5. januar 2004.
- Martin, RM, Smith, GD, Mangtani, P., Frankel, S. & Gunnell, D. (2002) Assosiasjon mellom amming og vekst: Boyd-Orr-kohortestudien. Archives of Diseases of Childhood - Fetal and Neonatal Edition 87, F193–201.
- Mayer Lawrence S. og McHugh Paul R., seksualitet og kjønn: Funn fra biologiske, psykologiske og samfunnsvitenskapelige, The New Atlantis, Number 50, Fall 2016, p. 116. http://www.thenewatlantis.com/sexualityandgender
- Mbugua K. Seksuell orientering og hjernestrukturer: En kritisk gjennomgang av nyere forskning. Current Science Vol. 84, Nei. 2 (25 januar 2003), pp. 173-178 (6 sider). https://www.jstor.org/stable/24108095
- McConaghy, N., Hadzi-Pavlovic, D., Stevens, C., Manicavasagar, V., Buhrich, N. & Vollmer-Conner, U. (2006) Fødselsrekkefølge og forholdet mellom heteroseksuelle / homoseksuelle følelser hos kvinner og menn ... Journal of Homosexuality 51, 161-174.
- McFadden Dennis og Shubel Erin, “Relative lengder på fingre og tær hos menn og kvinner,” Hormoner og oppførsel 42, nr. 4 (2002): 492 - 500, http://dx.doi.org/10.1006/hbeh.2002.1833
- Milinski, M. (2006). Det viktigste histokompatibilitetskomplekset, seksuell seleksjon og kameratvalg. Årlig gjennomgang av økologi og systematikk, 37, 159 - 186.
- Mitter C, Jakab A, Brugger PC, et al. Validering av in-utero-traktografi av humane fosterkommisjonelle og indre kapselfibre med histologisk struktur Tensoranalyse. Frontiers in Neuroanatomy. 2015; 9: 164. doi: 10.3389 / fnana.2015.00164.
- Morikawa, M., Yamada, H., Kato, EH, Shimada, S., Yamada, T. & Minakami, H. (2004) Embryo tap mønster er dominerende i spontanaborter med normal kromosom karyotype blant kvinner med gjentatt abort. Menneskelig reproduksjon 19, 2644-2647.
- Mukherjee, Siddhartha. Genen: En intim historie. Simon og Schuster, New York, 2016.
- Mustanski BS, Dupree MG, Nievergelt CM, Bocklandt S, Schork NJ, Hamer DH. En genombrannskanning av mannlig seksuell legning. Hum Genet 2005 Mar; 116 (4): 272-8. Epub 2005 Jan 12.
- New York Native, 7-10-1995, Gay Gene-forskningen holder ikke under granskning, Chicago Tribunes John Crewdson avdekker mulig vitenskapelig misbruk av NCI Researcher.
- NewsBeat (2015) Ugjendrivelig og vitenskapelig bevis Michelle Obama er faktisk en mann ... NewsBeat Ent. 24.11.2015. newsbeat.co.ke/gossip/irrefutable-scientific-proof-michelle-obama-is-indeed-a-man/
- Newsweek: februar 24, 1992 s.49
- NIAAA (2012) En familiehistorie om alkoholisme. Nasjonalt institutt for alkoholmisbruk og alkoholisme. https://pubs.niaaa.nih.gov/publications/familyhistory/famhist.htm
- Nimmons D. Sex and the Brain. Discover. 01.03.1994. discovermagazine.com/1994/mar/sexandthebrain346
- Ngun TC, Guo W, Ghahramani NM, Purkayastha K, Conn D, Sanchez FJ, Bocklandt S, Zhang M, Ramirez CM, Pellegrini M, Vilain E. En ny prediktiv modell for seksuell orientering ved bruk av epigenetiske markører. Sammendrag: En ny prediktiv modell av seksuell orientering ved bruk av epigenetiske markører. Presentert på American Society of Human Genetics 2015 årsmøte. Baltimore, Md.
- Nokia MS et al. Fysisk trening øker voksen hippocampal neurogenese hos hannrotter forutsatt at den er aerob og vedvarende. J Physiol. 2016 Apr 1; 594 (7): 1855-73. doi: 10.1113 / JP271552. Epub 2016 Feb 24.
- Norton R. Er homofili arvet? New York Review of Books, (juli, 1995). www.pbs.org/wgbh/pages/frontline/shows/assault/genetics/nyreview.html
- Nunez, JL & McCarthy, MM (2003) Kjønnsforskjeller og hormonelle effekter i en modell av for tidlig spedbarns hjerneskade. Annaler fra New York Academy of Sciences 1008, 281-284.
- Paglia C. Vamps & Tramps: New Essays. Vintage Books, 1994, på s. 71-72
- Parshley Lois. Kan genene dine få deg til å drepe? Populærvitenskap. 28.04.2016. https://www.popsci.com/can-your-genes-make-you-kill
- Paul, JP, et al. (2001) Å forstå seksuelle overgrep mot barn som en prediktor for seksuell risikotaking blant menn som har sex med menn: Urban Men's Health Study. Overgrep mot barn Negl. 25, 557 - 584.
- Paulhus, DL (2008) .Birthorder.InM.Haith (Red.), Oppslagsverk for spedbarns- og barndomsutvikling (Vol. 1, s. 204 - 211). San Diego, CA: AcademicPress. https://doi.org/10.13140/2.1.3578.3687.
- Paus T. Kartlegging av hjernemodning og kognitiv utvikling i ungdomsårene. Trender i kognitive vitenskaper. 2005; 9 (2): 60-68. https://doi.org/10.1016/j.tics.2004.12.008
- Pierik, FH, Burdorf, A., Deddens, JA, Juttmann, RE, & Weber, RFA (2004). Maternelle og farlige risikofaktorer for kryptorkisme og hypospadi: En case-control studie hos nyfødte gutter. Miljø- og helseperspektiver, 112, 1570-1576
- Poasa, KH, Blanchard, R., & Zucker, KJ (2004). Fødselsrekkefølge hos transkjønnede menn fra Polynesia: En kvantitativ studie av samoansk fa'-afafine. Journal of Sex and Marital Therapy, 30, 13-23. doi: 10.1080 / 00926230490247110.
- Pulst SM .. Genetisk koblingsanalyse. Arch Neurol. 1999; 56 (6): 667 - 672. doi: 10.1001 / archneur.56.6.667
- Pumberger, W., Pomberger, G. & Geissler, W. (2001) Proctocolitis hos ammende spedbarn: et bidrag til differensialdiagnose av hematochezia i tidlig barndom. Postgraduate Medical Journal 77, 252-254.
- Purcell, DW, Blanchard, R., & Zucker, KJ (2000). Fødselsrekkefølge i et moderne utvalg av homofile menn. Archives of Sexual Behavior, 29, 349–356.
- Queer etter valg. Gayle Madwin http://www.queerbychoice.com/
- Rahman Qazi og Wilson Glenn D., "Seksuell orientering og 2nd til 4th fingerlengdes forhold: bevis for organisering av effekter av kjønnshormoner eller utviklingsmessig ustabilitet?", Psychoneuroendocrinology 28, nr. 3 (2003): 288 - 303, http://dx.doi.org/10.1016/S0306-4530(02)00022-7
- Rainer, J. D., Mesnikoff, A., Kolb, L. C., Carr, A., "Homosexuality and Heterosexuality in Identical Twins" (inkludert en diskusjon av F. J. Kallmann) Psychosom Med. 1960;22:251
- Ramagopalan SV, Dyment DA, Handunnetthi L, Rice GP, Ebers GC. En genomomfattende skanning av mannlig seksuell legning. J Hum Genet. 2010 Feb; 55 (2): 131-2. http://dx.doi.org/10.1038/jhg.2009.135
- Remafedi G, et al. (1992) Demografi av seksuell tiltrekning hos ungdom. Pediatri 89, 714 - 721.
- Rice G et al., "Mannlig homoseksualitet: Fravær av kobling til mikrosatellittmarkører ved Xq28," Science 284, nr. 5414 (1999): 665 - 667, http://dx.doi.org/10.1126/science.284.5414.665
- Richiardi, L., Akre, O., Lambe, M., Granath, F., Montgomery, SM & Ekbom, A. (2004) Fødselsrekkefølge, søskipsstørrelse og risiko for bakteriekreft testikkelkreft. Epidemiologi 15, 323-329.
- Rind, B. (2001) Homofile og bifile tenåringsgutters seksuelle opplevelser med menn: en empirisk undersøkelse av psykologiske korrelater i en ikke-klinisk prøve. Arkiv for seksuell atferd 30, 345 - 368.
- Risch N, Squires-Wheeler E, Keats BJ. Mannlig seksuell legning og genetisk bevis. Science. 1993 desember 24; 262 (5142): 2063-5. DOI: 10.1126 / science.8266107
- Robinson SJ og Manning John T., “Forholdet mellom 2nd og 4th sifferlengde og mannlig homoseksualitet,” Evolution and Human Behaviour 21, nr. 5 (2000): 333 - 345, http://dx.doi.org/10.1016/S1090-5138(00)00052-0
- Rohrer, JM, Egloff, B., & Schmukle, SC (2015). Undersøkelse av fødselsordens innvirkning på personlighet. Proceedings of the National Academy of Sciences, 112,14224-14229. Https://doi.org/10.1073/pnas.1506451112.
- Rosario & Scrimshaw 2014, APA-håndbok, bind 1, s. 579
- Rosenthal, D., "Genetisk teori og unormal atferd" 1970, New York: McGrawHil
- Salg A, et al. Miljø og hjerneplastisitet: Mot en endogen farmakoterapi. Psychologica anmeldelser 2014; Vol. 94, Nei. 1. https://doi.org/10.1152/physrev.00036.2012
- Laks, C. (2012). Fødselsrekkefølge, effekt på personlighet og atferd. I V. Ramachandran (Red.), Encyclopedia of human behavior (Vol. 1, s. 353 - 359). London: Elsevier. https://doi.org/10.1016/B978-0-12-3750 00-6.00064-1.
- Sandberg, DE, Meyer-Bahlburg, HFL, Yager, TJ, Hensle, TW, Levitt, SB, Kogan, SJ & Reda, EF (1995) Kjønnsutvikling hos gutter født med hypospadier. Psychoneuroendocrinology 20, 693–709
- Sanders AR et al., "Genomfattende skanning demonstrerer betydelig kobling for mannlig seksuell legning," Psykologisk medisin 45, nr. 07 (2015): 1379 - 1388, http://dx.doi.org/10.1017/S0033291714002451
- Sanders AR, et al. Genome-Wide Association Study of Male Sexual Orientation. Sci rep. 2017; 7: 16950. http://dx.doi.org/10.1038/s41598-017-15736-4
- Satinover J. Homoseksualitet og sannhetens politikk. Raker Books 1996.
- Savic I, et al., "Hjernerespons på formodede feromoner hos homoseksuelle menn," Proceedings of the National Academy of Sciences 102, nr. 20 (2005): 7356 - 7361, http://dx.doi.org/10.1073/pnas.0407998102
- Savin-Williams, R.C & Ream, GL (2006) Pubertetsutbrudd og seksuell orientering i et ungdoms nasjonalt sannsynlighetsutvalg. Arkiv for seksuell atferd 35, 279-286.
- Science Media Center (2015). Ekspertreaksjon på konferansepresentasjon (upublisert arbeid) om epigenetikk og mannlig seksuell orientering. Oktober 8, 2015. http://www.sciencemediacentre.org/expert-reaction-to-conference-presentation-unpublished-work-on-epigenetics-and-male-sexual-orientation/
- Semenyna, SW, Petterson, LJ, VanderLaan, DP, & Vasey, PL (2017). En sammenligning av reproduksjonseffekten blant slektningene til samoanske androfile fa'afafine og gynefile menn. Archives of Sexual Behavior, 46, 87–93.
- Serano, J. M. (2010). "Saken mot autogynefili." International Journal of Transgenderism. 12 (3): 176–187. doi:10.1080/15532739.2010.514223
- Smith, MJ, Creary, MR, Clarke, A. & Upadhyaya, M. (1998) Kjønnsforhold og fravær av uniparental disomi ved spontane aborter med en normal karyotype. Klinisk genetikk 53, 258-261.
- Sorensen, HT, Olsen, ML, Mellemkjaer, L., Lagiou, P., Olsen, JH & Olsen, J. (2005) Den intrauterine opprinnelsen til brystkreft hos menn; en fødselsordrestudie i Danmark. European Journal of Cancer Prevention 14, 185-186.
- Speiser PW et al., "Medfødt binyrehyperplasi på grunn av steroid 21-hydroksylase-mangel: En endokrin samfunns klinisk praksisretningslinje," Journal of Clinical Endocrinology and Metabolism 95, nr. 9 (2009): 4133 - 4160, http://dx.doi.org/10.1210/jc.2009-2631
- Speiser PW, White PC, “Medfødt binyrehyperplasi”, New England Journal of Medicine 349, nr. 8 (2003): 776 - 788, http://dx.doi.org/10.1056/NEJMra021561
- Stein, Edward, The Mismeasure of Desire: The Science, Theory and Ethics of Sexual Orientation (New York: Oxford University Press, 1999), 145
- Sulloway, FJ (1996). Født til opprør: Fødselsrekkefølge, familiedynamikk og kreative liv. New York: Pantheon Books.
- Swaab DF, "Seksuell orientering og dens grunnlag i hjernestruktur og funksjon," Proceedings of the National Academy of Sciences 105, nr. 30 (2008): 10273 - 10274, http://dx.doi.org/10.1073/pnas.0805542105
- Tannehill B. New Yorker siterer skammelig Anti-LGBT 'forsker'. Bilerico-prosjektet. 29. juli 2014. bilerico.lgbtqnation.com/2014/07/new_yorker_shamefully_cites_anti-lgbt_researcher.php
- Taylor, Tim, "Tvillingstudier av homoseksualitet," Avhandling, Institutt for eksperimentell psykologi, University of Cambridge, 1992.
- The New York Times (2004). Bryllup / feiring; Dean Hamer, Joseph Wilson. April 11, 2004. www.nytimes.com/2004/04/11/style/weddings-celebrations-dean-hamer-joseph-wilson.html (01.12.2017 bekreftet)
- Restitusjonsbyen (2017). Hvorfor alkoholisme ikke er arvelig. Restitusjonsbyen. https://www.therecoveryvillage.com/alcohol-abuse/faq/alcoholism-not-hereditary/#gref
- Theodosis DT, et al. Aktivitetsavhengig neuronal glial og synaptisk plastisitet i voksen pattedyrhypothalamus. Neuroscience Volume 57, Issue 3, December 1993, Pages 501-535. https://doi.org/10.1016/0306-4522(93)90002-W
- Tomeo, ME, Templer, DI, Anderson, S. & Kotler, D. (2001) Sammenligningsdata for overgrep fra barndom og ungdom hos heterofile og homofile personer. Arkiv for seksuell atferd 30, 535-541.
- Tsroadsmap. Kategorisk galt? A Bailey-Blanchard-Lawrence clearinghousehttp: //www.tsroadmap.com/info/bailey-blanchard-lawrence.html
- Turner, MC, Bessos, H., Fagge, T., Harkness, M., Rentoul, R., Seymour, J. et al. (2005) Prospektiv epidemiologisk studie av utfallet og kostnadseffektiviteten av antenatal screening for å oppdage neonatal alloimmun trombocytopeni på grunn av ani-HPA-1a. Transfusjon 45, 1945 - 1956.
- Rahman Q. http://dx.doi.org/10.1037/0735-7044.117.5.1096
- Van Ombergen, A., Jillings, S., Jeurissen, B., Tomilovskaya, E., Rühl, RM, Rumshiskaya, A., ... Wuyts, FL (2018). Hjernevev - Volumendringer i kosmonauter. New England Journal of Medicine, 379 (17), 1678 - 1680. doi: 10.1056 / nejmc1809011
- VanderLaan, DP, Blanchard, R., Wood, H., Garzon, LC, & Zucker, KJ (2015). Fødselsvekt og to mulige typer maternelle effekter på mannlig seksuell legning: En klinisk studie av barn og ungdom henvist til en kjønnsidentitetstjeneste. Utviklingspsykobiologi, 57,25-34. https://doi.org/10.1002/dev.21254.
- Voracek Martin, Manning John T., og Ponocny Ivo, “sifferforhold (2D: 4D) hos homoseksuelle og heterofile menn fra Østerrike,” Archives of Sexual Behaviour 34, nr. 3 (2005): 335 - 340, http://dx.doi.org/10.1007/s10508-005-3122-x
- Wedekind, C., Seebeck, T., Bettens, F., & Paepke, AJ (1995). MHC-avhengige kameratpreferanser hos mennesker. Proceedings Biological Sciences, 22, 245-249.
- Wellings, K, et al. (1994) Seksuell atferd i Storbritannia: The National Survey of Sexual Attitudes and Lifestyles. Penguin Books, London
- Whitehead NE. Et antistoffantistoff? Re-undersøkelse av immunhypotesen til mors. J. biosoc. Sci. 2007. doi: 10.1017 / S0021932007001903
- Williams TJ et al., "Fingerlengdeforhold og seksuell legning," Nature 404, nr. 6777 (2000): 455 - 456, http://dx.doi.org/10.1038/35006555
- Williams, Zev (20. september 2012). "Indusere toleranse for graviditet." New England Journal of Medicine. 367:1159–1161. doi:10.1056/NEJMcibr1207279. PMC 3644969
- Wilson JD, et al. Den hormonelle kontrollen av seksuell utvikling. Science 211 (1981): 1278 - 1284, http://dx.doi.org/10.1126/science.7010602
- Witchel Alex. Intervju med Cynthia Nixon. New York Times Magazine. Livet etter "Sex". Januar 2012. http://www.nytimes.com/2012/01/22/magazine/cynthia-nixon-wit.html
- Wyndzen, M. H. (2003). Autogynephilia og Ray Blanchards feilstyrte sexlystmodell for transseksualitet. Alt blandet sammen: En transkjønnet psykologiprofessors perspektiv på livet, kjønnspsykologien og "kjønnsidentitetsforstyrrelse". Tilgjengelig: http://www.GenderPsychology.org/autogynpehilia/ray_blanchard/
- Wyre, R. (1990) Hvorfor misbruker menn seksuelt barn? I Tate, T. (red.) Barnepornografi. Methuen, London, pp. 281 - 288.
- Xanthakos, SA, Schwimmer, JB, Aldana, HM, Rothenberg, ME, Witte, DP & Cohen, MB (2005) Prevalens og utfall av allergisk kolitt hos friske spedbarn med rektal blødning: en potensiell kohortestudie. Journal of Pediatric Gastroenterology and Nutrition 41, 16-22.
- Yong E. Nei, forskere har ikke funnet den homofile genen. Mediene hyper på en studie som ikke gjør som den sier at den gjør. Science. Oktober 10, 2015. https://www.theatlantic.com/science/archive/2015/10/no-scientists-have-not-found-the-gay-gene/410059/
- Zanin E, Ranjeva JP, Confort-Gouny S, et al. Modning av hvit substans av normal menneskelig føtale hjerne. En in vivo diffusjons-tensortraktografistudie. Hjerne og atferd. 2011; 1 (2): 95-108. doi: 10.1002 / brb3.17.
- Zietsch BP. Årsaker til forsiktighet angående den fødselsmessige fødselsorden. Arch Sex Behav. 2018. DOI 10.1007 / s10508-017-1086-2
- Zietsch, BP, Verweij, KJH, Heath, AC, Madden, PAF, Martin, NG, Nelson, EC, ... Lynskey, MT (2012). Bidrar delte etiologiske faktorer til forholdet mellom seksuell legning og depresjon? Psykologisk medisin, 42,521 - 532. doi: 10.1017 / s0033291711001577
- Zusman, I., Gurevich, P. & Ben-Hur, H. (2005) To sekretoriske immunsystemer (slimhinne og barriere) i human intrauterin utvikling, normal og patologisk (Review). International Journal of Molecular Medicine 16, 127-133.
Selv om man tar hensyn til eneggede tvillinger, er homofili justert 1:1. Og så er det nødvendig å gi foreldre sykelighet, økonomiske problemer med å opprettholde kvaliteten på helsen og sikre de samme kontaktene, familieproblemer, kriminelle risikoer og så videre, som barnet deres vil bli utsatt for, hvis lykke alle bryr seg så mye om , invitere ham til selvstendig (?) å velge en slik livsstil . Jeg prøver å gjøre dette, men de begynte å blokkere meg.
En fornuftig person tror jeg forstår at dette er bedriftens interesse. for å si det mildt. Som spesialist i den føderale tjenesten for menneskelig velvære, anbefaler jeg oppriktig ikke slik lykke, som ikke "lukter" av ikke bare lykke, men også en oppblåst standard for velvære. Jeg kan ikke forestille meg at noen kan utvikle noen hygieniske sikkerhetsanbefalinger for denne typen sex (spøk med tårer...). Jeg skal forresten prøve å lete etter det.